в оглавление
назад
Клетки и ткани
Клетки представляют собой основные единицы живого, из которых строится весь организм. Функционирование тех или иных органов зависит прежде всего от активности клеток, из которых эти органы образованы. В свою очередь почти все системы органов прямо или косвенно поставляют клеткам материалы, необходимые для их жизнедеятельности, и создают среду, необходимую для их существования. Каждая клетка живет собственной жизнью, но продолжительность ее жизни зависит от функционирования других клеток, и каждая клетка в свою очередь вносит вклад в благополучие организма в целом.
Строение и функции клетки
Клетки тела чрезвычайно многообразны по строению и функциям. Но несмотря на структурные и функциональные различия в строении клеток по крайней мере высших животных, к которым принадлежат и млекопитающие, имеются общие черты.

Схема строения типичной животной клетки: 1 — цитоплазматическая мембрана; 2 — пиноцитозные пузырьки; 3 — центросома; 4 — гиалоплазма; 5 — зернистая эндоплазматическая сеть: а — мембрана зернистой сети, б — рибосомы; 6 — связь перинуклеарного пространства с полостями эндоплазматической сети; 7 — ядро; 8 — ядерная пора; 9 — незернистая (гладкая) эндоплазматическая сеть; 10 — ядрышко; 11 — комплекс Гольджи; 12 — секреторные вакуоли; 13 — митохондрия; 14 — лизосомы; 15 — три последовательные стадии фагоцитоза; 16 — связь клеточной оболочки с мембранами.
В центре клетки обычно располагается ядро. В нем находится большое количество вещества хроматина, представляющего собой дезоксирибонуклеиновую кислоту (ДНК). Перед началом клеточного деления хроматин уплотняется, образуя ряд парных палочковидных структур — хромосом. В процессе деления каждая хромосома расщепляется в продольном направлении, поэтому дочерние клетки получают по полному набору хромосом, а следовательно, и содержащегося в них генетического материала. Другое вещество — рибонуклеиновая кислота (РНК) — образует компактную структуру, называемую ядрышком. За исключением того времени, когда клетка делится, ядро отделено от остального содержимого клетки мембраной. В этой мембране имеются т. н. «поры», которые на самом деле представляют собой скорее плотные тяжи, а не отверстия, как можно было бы ожидать, исходя из их названия. Снаружи клетка ограничена ясно выраженной клеточной мембраной, которая в значительной степени регулирует поступление в клетку различных веществ и их выведение. Мембрана, в которой можно видеть три слоя, образована липидами и белками. Содержимое клетки в целом называют протоплазмой; цитоплазма — более узкий термин, обозначающий содержимое клетки без ядра.
В цитоплазме находится ряд дискретных структур, называемых органеллами. Вблизи ядра лежат маленькие тельца — центриоли, участвующие в клеточном делении. По цитоплазме рассеяны многочисленные мелкие частицы — митохондрии. Это вытянутые сфероиды с рядом внутренних поперечных перегородок, называемых кристами. В митохондриях содержится большое количество ферментов, катализирующих химические реакции, благодаря которым клетка получает необходимую ей энергию. Именно в митохондриях происходит окисление сахаров и жирных кислот, в результате чего образуется аденозинтрифосфат (АТФ) — универсальный носитель энергии в клетке. Митохондрии в отличие от других органелл цитоплазмы содержат также некоторое количество дезоксирибонуклеиновой кислоты и частично независимы от ядра. Во многих клетках имеется аппарат Гольджи, нередко называемый также «сетчатым (ретикулярным) аппаратом». Он состоит из стопок уплощенных блюдцевидных структур (цистерн) и мелких пузырьков, функция которых заключается в накоплении белков, к которым присоединяются вырабатываемые здесь крупные углеводные группы, после чего эти белковоуглеводные комплексы выделяются из клетки в виде секреторных гранул. От поверхности структур аппарата Гольджи могут отпочковываться лизосомы — мембранные пузырьки, содержащие гидролитические ферменты.
Долгое время полагали, что, не считая только что описанных структур, цитоплазма представляет собой, в сущности, аморфный желеобразный коллоид, где в жидкой водянистой среде растворены органические вещества. Однако было выявлено, что цитоплазма обладает ультраструктурой: вся она пронизана сетью тонких мембран или трубочек, образующих т. н. эндоплазматический ретикулум, а пространство между ними заполнено жидкой средой. Цитоплазма содержит также многочисленные маленькие частицы, называемые рибосомами, которые встроены в эти мембраны и не видны под обычным микроскопом; рибосомы служат основными центрами синтеза белка. В этом случае эндоплазматический ретикулум называют шероховатым или гранулярным; частично его мембраны могут принимать участие в формировании аппарата Гольджи. Если эндоплазматический ретикулум не содержит рибосом, то его называют гладким или агранулярным; агранулярный эндоплазматический ретикулум участвует в синтезе липидов (в том числе стероидных гормонов) и в регуляции концентрации ионов кальция (особенно в саркоплазматическом ретикулуме мышечных клеток).
Клетка может содержать также различные другие включения или тельца. Разумеется, в ней имеются секреторные гранулы, но, кроме того, вакуоли с поглощенными веществами, пигментные гранулы и жировые капли.
Поверхность клетки не менее сложна, чем ее содержимое. Соседние клетки бывают «связаны» специальными структурами: десмосомы просто удерживают две клетки вместе, тогда как щелевые контакты участвуют в переносе веществ между этими клетками. Свободные поверхности иногда несут щеточную каемку, состоящую из множества микроворсинок (каждая ворсинка тонкого кишечника состоит из многих клеток, которые могут быть в свою очередь покрыты микроворсинками). Реснички представляют собой более крупные и более сложные нитевидные подвижные структуры.
Такая сложность структуры клетки создает впечатление статичности ее со стояния. На самом деле все обстоит как раз наоборот. Каждая клетка должна производить энергию для выполнения своей функции в организме (будь то секреция, проведение нервных импульсов, мышечная работа или что-то иное); однако, кроме того, каждая клетка непрерывно претерпевает химические изменения, необходимые для поддержания своего собственного существования. Клетка, подобно организму в целом, обладает, так сказать, собственным «основным обменом», обеспечивающим сохранение ее жизненных функций.
Клеточная среда и тканевая жидкость
Любой организм должен создавать физическую и химическую среду, удовлетворяющую потребностям его клеточных единиц. Для клеток позвоночных эти потребности определены достаточно строго.
Организм как целое может существовать в довольно широком диапазоне температур внешней среды, но колебания температуры тела, которые могут выдержать его клетки, ограниченны. Ниже допустимого предела организм гибнет от переохлаждения; при выходе за верхнюю планку температурного диапазона наступает смерть от перегрева, так как некоторые белки претерпевают необратимую денатурацию.
Клетка в состоянии жить и избегать высыхания, только находясь в жидкой среде. Такой средой служит тканевая жидкость (ее называют также интерстициальной или межклеточной жидкостью), пропитывающая все ткани тела. Воды как таковой клеткам недостаточно. Тканевая жидкость должна содержать довольно значительное количество растворенных в ней веществ; в противном случае осмотическое давление может вызвать разбухание и разрыв омываемых ею клеток. Кроме того, клетки лучше всего себя чувствуют как раз при том составе растворенных веществ (это главным образом соли неорганических кислот), который и характерен для обычной тканевой жидкости. Последняя в значительном количестве содержит ионы натрия и хлора, в меньших — ионы калия, кальция и магния, а также небольшие количества других элементов.
Почему клеткам необходим именно такой состав тканевой жидкости (отличающийся от состава внутренней среды самой клетки), остается неясным. Интересно, однако, отметить, что за двумя исключениями (отсутствие сульфатов и более низкое содержание магния) тканевая жидкость позвоночных сходна по составу с морской водой, хотя обычно сильнее разбавлена. Возможно, это не случайно. Предки позвоночных были животными, организованными весьма просто, омывавшимися и пропитанными водами раннепалеозойского океана; морская вода была главным фактором среды, в которой протекала их эволюция. Будучи уже сложными организмами, они независимо создали себе внутреннюю среду: содержащую соли интерстициальную жидкость, которая явилась, если можно так выразиться, остатком древнего моря. Тканевая жидкость относительно свободно сообщается со сходной жидкостью, образующей плазму крови, так что равномерное распределение жидкостей в организме обеспечивается системой кровообращения.
Большую часть веществ, необходимых клетке для существования, она получает из омывающей ее тканевой жидкости. В эту же жидкость выделяет конечные продукты обмена. Тесный контакт тканевой жидкости с системой кровообращения предоставляет возможность непрерывно получать питательные вещества и освобождаться от конечных продуктов обмена.
Природа клеточной мембраны и необходимость наличия в тканевой жидкости достаточной концентрации растворенных веществ, для того чтобы уравновешивать их концентрацию в клетке, запускают физико-биологический механизм осмоса. Если два объема жидкости (например, воды с растворенными в ней веществами) разделены мембраной, искусственной или естественной, то мембрана может быть либо настолько непроницаемой, что обмен веществами через нее почти или совсем невозможен, или же такой проницаемой, что через нее будут легко проходить все растворенные в жидкостях вещества, т. е. будет происходить свободный обмен. Существуют, однако, мембраны промежуточного типа, не пропускающие большие молекулы или даже большие ионы, хотя более мелкие легко через них проходят. Такие мембраны называют полупроницаемыми. Клеточные мембраны различной степени проницаемости имеются в клетках многих органов. Очевидно, что осмос — процесс, играющий заметную роль в определении структуры тела. Следует помнить, что это совершенно иной процесс, нежели активный перенос определенных веществ через клеточные мембраны, часто происходящий во многих тканях.
Эпителий
Клетки тела не являются обособленными единицами: они входят в состав тканей — организованных ассоциаций клеток одинакового происхождения, выполняющих сходные функции. В некоторых случаях, особенно в случае соединительной ткани (в широком смысле), агрегация клеток может быть относительно диффузной, а в других — слабо выраженной или аморфной. В самом деле, довольно трудно освоиться с мыслью, что клетки циркулирующей крови образуют некую ткань. Однако часто ассоциация клеток образует такую ткань, как эпителий — упорядоченный и плотный пласт клеток, который обычно одной своей стороной ограничивает поверхность тела или одну из его внутренних полостей.
У зародыша эпителий почти всех типов имеет довольно простую и четкую структуру. В дальнейшем, однако, в результате утолщения и модификации некоторые типы в значительной степени утрачивают сходство с эпителием, как, например, в толще печени. Эта дифференцировка эпителия в процессе онтогенеза примерно соответствует его вероятной дифференцировке в процессе филогенеза. У самых первых и самых простых многоклеточных организмов предположительно имелся эпителий лишь одного типа. Он покрывал их тело снаружи и выполнял самые разнообразные функции: защитную, восприятие раздражений, поглощение питательных веществ из внешнего мира и выделение в среду конечных продуктов жизнедеятельности. В дальнейшем, с образованием внутренних полостей и внутренних структур, стала не только возможной, но и необходимой специализация эпителиальных тканей. Следует отметить, что в значительной части пищеварительного тракта и в других внутренних органах эпителий, защищенный от вредных внешних воздействий, навсегда остается тонким и нежным, а его клетки могут обладать такими важными свойствами, как способность к всасыванию или к секреции. Такой эпителий обычно называют слизистой оболочкой, так как некоторые или все его клетки секретируют слизь; в результате этот эпителий всегда покрыт тонкой пленкой жидкости, которая увлажняет его.

Строение покровного эпителия: А — простой плоский эпителий; Б — простой кубический эпителий; В — простой столбчатый эпителий; Г — реснитчатый эпителий; Д — переходный эпителий; Е — неороговевающий многослойный плоский эпителий.
Для удобства различные типы эпителиальных клеток называют по-разному. Плоские клетки очень низкие, их высота по сравнению с шириной очень мала, и на поперечных срезах они имеют вид тонких линий. У кубических клеток ширина примерно равна высоте, и на срезах они выглядят квадратными. Цилиндрические клетки больше в высоту, чем в ширину.
У развивающихся зародышей эпителий часто состоит из одного слоя клеток. Иногда он сохраняется в таком виде и во взрослом состоянии и известен под названием простого или однослойного эпителия. Такой эпителий встречается там, где опасность повреждения или снашивания невелика, или же там, где происходит всасывание или фильтрация. Однослойный эпителий может состоять из клеток любого из трех названных выше типов. Вначале клетки простого эпителия бывают округлыми, но по мере того как их упаковка становится плотнее, они обычно приобретают кубическую форму; эпителий такого типа довольно часто встречается у взрослых особей. Плоский эпителий характерен, например, для выстилки кровеносных сосудов и для других участков, где важную роль играет осмотический обмен через мембрану. Поверхностные клетки, несущие секреторные или другие существенные функции, обычно бывают более высокими, цилиндрическими.
Если эпителий состоит из двух или более слоев клеток, то его называют многослойным эпителием. Разным типам многослойного эпителия дают названия по характеру клеток, образующих поверхностный слой, т. е. многослойный эпителий может быть плоским, кубическим или цилиндрическим, хотя два последних типа встречаются редко. Более глубокие слои многослойного плоского эпителия могут состоять из кубических или цилиндрических клеток, причем нижележащие клетки цилиндрического типа могут быть сильно уплощенными.
Как правило, эпителиальные клетки имеют форму многоугольников, часто шестиугольников, что представляет собой оптимальный компромисс между идеально круглой формой каждой отдельной единицы и требованиями компактного расположения. В эпителиальной ткани соседние клетки могут быть плотно пригнаны одна к другой, но между ними все же имеются очень небольшие пространства, заполненные тканевой жидкостью. Эти пространства могут перекрываться «плотными контактами» между соседними клетками; такие контакты, по-видимому, ограничивают передвижение веществ в тканевой жидкости. Поскольку плотные контакты обычно относительно непроницаемы, они не допускают переноса веществ из клетки в клетку, который происходит, как отмечалось ранее, в области щелевых контактов — мельчайших пор, пронизывающих мембраны соседних клеток. Как правило, в эпителии нет кровеносных сосудов, и питательные вещества проникают в клетки, поднимаясь вверх с интерстициальной жидкостью из сосудов, расположенных под основанием эпителия. В большинстве случаев эпителий снизу подстилает соединительная ткань; между двумя тканями обычно лежит базальная мембрана, образуемая главным образом или полностью эпителиальными клетками (базальная пластинка), а иногда также соединительной тканью (ретикулярная пластинка).
Для многослойного эпителия часто характерно постоянное изнашивание и слущивание клеток, образующих поверхностный слой. Утраченные клетки замещаются снизу, где клетки эпителия образуют герминативный слой, способный обеспечить замещение за счет клеточного деления. Главные функции эпителия в целом связаны с его свободной наружной поверхностью; поэтому неудивительно, что во многих типах многослойного эпителия по мере приближения к поверхности специализация клеток возрастает. Кроме того, в эпителии почти любого типа клетки более или менее поляризованы: проксимальный (обращенный вглубь) и дистальный (обращенный к поверхности) концы клеток различаются по характеру.
На свободной поверхности эпителия часто имеются явные признаки специализации. Иногда клетки секретируют кутикулу, покрывающую эпителий более или менее плотным слоем. Чаще, однако, бывает изменена поверхность самих клеток. К числу наиболее специализированных структур принадлежат реснички, располагающиеся на поверхности цилиндрического эпителия различных типов. Это тонкие подвижные «органы», похожие на волоски и содержащие 11 микротрубочек, из которых две расположены в центре, а остальные 9 — по периферии; все микротрубочки прикреплены к базальному тельцу, имеющему сложную структуру. Реснички поочередно сгибаются, но лишь в одну сторону; в результате их движения (независимого от нервной регуляции) по покрытой ими поверхности как бы пробегает волна, непрерывно перемещая слизь или другие вещества в одном направлении. В некоторых случаях, однако, все реснички бьют в унисон.
Известны случаи, когда однослойный эпителий выглядит как многослойный; это т. н. псевдомногослойный (многорядный) эпителий. Так, если однослойный эпителий состоит из клеток двух типов, ядра которых расположены на разном уровне, или же из клеток разной высоты, то создается впечатление, что в нем имеются два слоя. Термин «псевдомногослойный» более оправдан в тех случаях, когда на базальной мембране однослойного эпителия лежат клетки двух типов, но до поверхности доходят лишь клетки одного типа. Неудачно названный переходный эпителий, имеющийся в мочевом пузыре и мочеиспускательном канале, на самом деле представляет собой особый тип псевдомногослойного эпителия. Если полость, которую он выстилает, пуста или растянута не очень сильно, эпителий этого типа кажется очень многослойным; когда полость расширяется, то эпителий растягивается и соответственно утоньшается. Другие виды эпителия, например эпителий, выстилающий кровеносные сосуды, также должны растягиваться и сокращаться вместе с окружающими тканями.
В широком смысле эпителием можно назвать любую ткань, соответствующую приведенному описанию (т. е. пласты прилегающих друг к другу клеток), независимо от того, в какой части тела она находится. Однако, в большинстве случаев эпителием называют только слои ткани, покрывающие наружную поверхность тела, с одной стороны, и выстилающие внутреннюю поверхность кишечника и его производных — с другой, а также все полости, несомненно образованные этими наружными и внутренними поверхностями. Теоретической основой такого представления служит тот факт, что эпителиальные ткани образуются в процессе онтогенеза из покрытых эктодермой и энтодермой поверхностей зародыша. Если принять такое ограничение, то для тканей, выстилающих полости, которые образуются вторично в более глубоких участках организма, следует использовать один или несколько иных терминов. Эндотелием обычно называют эпителиальную выстилку кровеносных сосудов. Иногда этот термин используют также для обозначения выстилки полостей тела, однако эту выстилку предпочтительнее называть мезотелием.
Железы
Секреторная активность — синтез и выделение жидких веществ — свойственна ряду различных клеток; клетки, для которых это основной вид активности, называют железистыми клетками.
Железы почти всегда имеют эпителиальное происхождение, хотя нервная и другие ткани также могут обладать способностью к секреции. Нередко железы сразу различимы на гистологических препаратах благодаря наличию в их клетках гранул секретируемых веществ или вакуолей, наполненных секретом. Железы и железистые клетки можно разделить на группы, исходя из тех последствий для клетки, которыми сопровождается выделение секретируемых веществ. Если секреторная активность происходит постоянно, не причиняя клетке видимого вреда, то железу называют мерокриновой. Но если секретируемое вещество густое или вязкое, клетки при его выделении обычно повреждаются. В апокриновых железах при выделении секрета разрушается только верхушечная часть клетки, но затем она восстанавливается и секреция продолжается. Таковы, например, потовые железы. Однако в некоторых железах клетка при выделении своего содержимого разрушается полностью; такие железы называют голокриновыми. К этой категории относят, к примеру, сальные железы кожи.
Во многих случаях железистые клетки встречаются поодиночке или небольшими группами на ограниченных участках эпителия, выполняющего также и другие функции. Слизь — густая вязкая смазка, увлажняющая и защищающая различные мембраны,— вырабатывается особыми железами, но нередко ее выделяют отдельные клетки, разбросанные по поверхности эпителия. Секрет часто накапливается в верхних частях клеток в виде крупных капель, растягивая клетки, которые по этой причине получили название бокаловидных.
Хотя железистые клетки или железистый участок могут просто составлять часть поверхности эпителия, нередко в процессе эмбрионального развития железистые клетки обособляются от общей эпителиальной поверхности и образуют дискретные железы. В большинстве случаев секрет желез выводится через протоки в какую-либо полость тела или на его поверхность; такие железы называют экзокринными. В результате обособления железы от эпителия решается ряд проблем. Благодаря складчатости железистого эпителия сильно увеличивается секреторная поверхность; железистые клетки получают защиту от различных опасностей, которым может подвергнуться эпителиальная поверхность; наличие узкого протока (образующие его клетки обычно не бывают секреторными) делает возможным регуляцию выделения секрета; создаются условия для накопления значительных количеств секрета.
Экзокринные железы весьма разнообразны. Секреторные участки имеют форму трубочек или округлых карманов (альвеолы или ацинусы) и бывают простыми или сложными. Встречаются также железы промежуточного типа (трубчато-альвеолярные и т. п.). Кроме того, в крупных железах иногда разветвляются и протоки, что приводит к разделению железы на доли и дольки. Железы часто имеют очень неправильную форму, поскольку главную роль играет их объем, а форма при этом несущественна. Крупные железы обычно окружены соединительной тканью, которая заходит также в промежутки между долями. Обычно железы обильно снабжены капиллярами и нервными окончаниями.
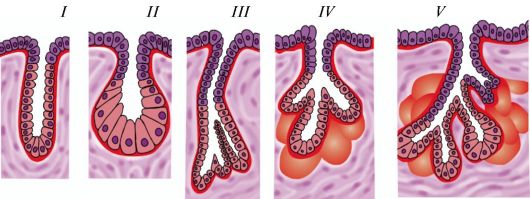
Типы экзокринных желез: I — простая трубчатая железа с неразветвленным начальным отделом; II — простая альвеолярная железа с неразветвленным начальным отделом; III — простая трубчатая железа с разветвленным начальным отделом; IV — простая альвеолярная железа с разветвленным начальным отделом; V — сложная альвеолярно-трубчатая железа с разветвленными начальными отделами.
Помимо «нормальных» экзокринных желез в организме имеются также эндокринные железы, выделяющие свой секрет не наружу, а в кровоток. В некоторых случаях можно проследить их образование из эпителия в процессе онтогенеза; возможно, что такие железы первоначально были экзокринными (например, в отношении щитовидной железы это, по-видимому, можно считать установленным). Что касается других эндокринных желез, то сведений о такой их родословной нет, и их филогенетическое происхождение остается весьма неопределенным. В самом деле, в некоторых случаях та или иная «железа» явно представляет собой нейтральную структуру. Поскольку секреты эндокринных желез, т. е. гормоны, переносимые с кровью, могут оказывать свое действие в любом участке организма, местоположение самих желез анатомически никак не связано с органами, на которые они действуют, и они могут находиться в самых разных областях тела. Как и в случае экзокринных желез, их форма никак не связана с их функцией и поэтому очень разнообразна.
Неэпителиальные ткани
Очень многие ткани, особенно выстилка пищеварительного тракта и ее производные, а также покровные ткани, сохраняют явно эпителиальный характер во взрослом организме. Другие ткани, происходящие из эпителия зародыша, у взрослой особи так сильно модифицированы, что их эпителиальная природа замаскирована. Например, печеночная ткань происходит из эпителия одного из карманообразных выростов кишечника, который превращается в желчные протоки взрослого организма. Но, хотя клетки печени взрослой особи сохраняют связь с ветвями желчного протока, массивная структура печеночной ткани мало похожа на первоначальную структуру эпителия. Большая часть нервной системы развивается из эпителия, образующего нервную трубку; хотя полости, соответствующие невроцелю (и выстилающие их эпителиальные клетки), сохраняются у взрослой особи, в нервной ткани почти нет следов структуры, типичной для эпителия. Можно считать, что из эпителия происходят и поперечнополосатые мышцы туловища. Но в поперечнополосатых мышцах взрослых животных не сохраняется никаких следов предковой эпителиальной структуры, и даже у зародыша признаки их эпителиального происхождения очень скоро исчезают.
Тем не менее, эти ткани в широком смысле можно рассматривать как эпителиальные по своей природе, поскольку они онтогенетически или филогенетически происходят из настоящего эпителия. Принято, однако, различать по крайней мере нервную и мышечную ткани как отдельные «основные типы», специализированные для проведения нервных импульсов и мышечного сокращения. Другие производные эпителия выделить не так просто, и поэтому их продолжают называть эпителиальными, несмотря на их модификации.
С некоторыми другими тканями — соединительной и скелетной, а также с тканями, образующими кровеносную систему, — дело обстоит иначе (клетки кровеносных сосудов могут, однако, быть эпителиальными (эндотелий) или мышечными (гладкие мышцы). Они развиваются из зародышевой ткани, известной под названием мезенхимы; эти клетки, возможно, и происходят из нижних поверхностей эпителия, но сами никогда не образуют компактную ткань эпителиального типа. Они распределены диффузно по основному веществу — матриксу, который сами и продуцируют. В скелетных элементах матрикс образует твердое вещество хряща или кости, в собственно соединительной ткани он студнеобразный, а в крови — жидкий (плазма). Все эти ткани можно рассматривать как соединительную ткань в широком смысле, которую принято считать четвертой из основных тканей (три другие — это эпителиальная, нервная и мышечная ткани).
Соединительные ткани
Соединительные ткани образуют «набивку» тела и армируют эпителии многих органов, обеспечивая телу прочность и опору. Простейший тип этих тканей — рыхлая (ареолярная) соединительная ткань. Значительная ее часть представлена студенистым белковым основным веществом. Оно содержит рыхлую сеть мелких ветвящихся ретикулиновых волокон и более многочисленных длинных и тонких белых коллагеновых волокон — гибких, но нерастяжимых структур, которые формируются веретеновидными или звездчатыми клетками соединительной ткани — фибробластами; каждое волокно состоит из пучка очень тонких фибрилл, которые образованы длинными молекулами белка коллагена.
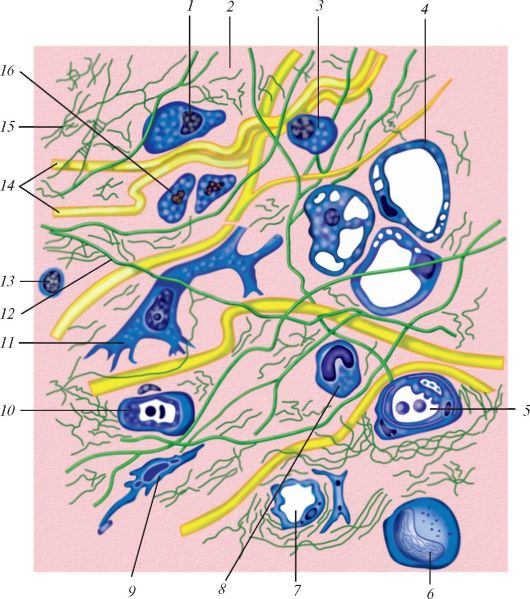
Схема строения рыхлой волокнистой соединительной ткани: 1 — макрофагоцит; 2 — аморфное межклеточное (основное) вещество; 3 — плазмоцит (плазматическая клетка); 4 — липоцит (жировая клетка); 5 — кровеносный сосуд; 6 — моноцит; 7 — лимфатический капиляр; 8 — эозинофильный гранулоцит; 9 — фиброцит; 10 — кровеносный капилляр; 11 — фибробласт; 12 — эластическое волокно; 13 — лимфоцит; 14 — коллагеновые волокна; 15 — ретикулярные волокна; 16 — тканевый базофил.
Другой распространенной разновидностью является компактная соединительная ткань; в ней плотно упакованная масса переплетенных волокон создает подобие войлока. Большинство видов соединительной ткани содержит небольшой процент желтых эластических волокон; в некоторых случаях эластические волокна преобладают, формируя эластичные ткани. Сухожилия (tendines), при помощи которых крепятся к скелету многие мышцы, состоят из параллельных пучков коллагеновых волокон; одним концом они крепко связаны с мышечными волокнами, другим прочно прикрепляются к хрящу или кости, внутрь которых могут проникать их волокна. Кроме того, некоторые очень подвижные сухожилия окружены волокнистой соединительнотканной оболочкой. Там, где сухожилия скользят по поверхности кости, под ними могут формироваться из соединительной ткани сумки (bursae) — наполненные жидкостью мешки. Связки (ligamenta) похожи на сухожилия, но соединяют между собой скелетные элементы; фасции (fasciae) — это листки соединительной ткани, одевающие мышцы или другие образования.
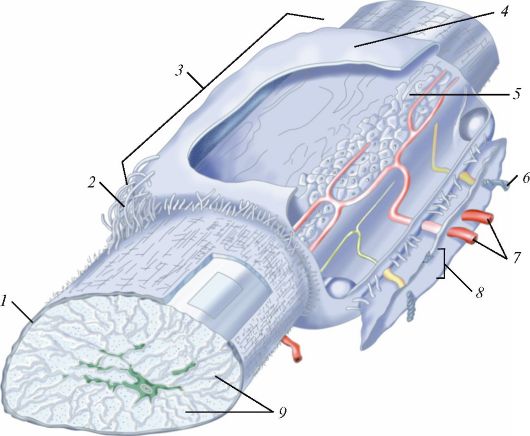
Схема строения сухожилия: 1 — перитендиний; 2 — фиброзный слой; 3 — сухожильное влагалище; 4 — париетальная часть влагалища; 5 — сухожильная часть влагалища; 6 — нервное волокно; 7 — кровеносные сосуды; 8 — мезотендрий; 9 — пучки сухожильных волокон I-го порядка.
Далее можно отметить присутствие в соединительных тканях тучных клеток, имеющих неправильную овальную форму и содержащих интенсивно окрашивающиеся гранулы. Они содержат запасы антикоагулянта гепарина, основного амина гистамина и сужающего сосуды серотонина; однако функция тучных клеток до конца еще не выяснена. Кроме того, в соединительной ткани обнаруживаются макрофаги (гистиоциты), играющие важную роль в иммунной защите организма. Могут также присутствовать форменные элементы крови, особенно лимфоциты.
Жировая, или адипозная, ткань тоже является разновидностью соединительной ткани; она образуется чаще всего под кожей или в складках брыжейки между органами брюшной полости. Внутри видоизмененных мезенхимных клеток формируются жировые капли; по мере образования мелкие капли сливаются в крупные, сильно растягивая эти клетки. Большая часть жира — беловатая, но иногда он бывает темнее. Существует два типа жировой ткани: белый жир с запасом триглицеридов и бурый жир, состоящий из клеток с большим количеством митохондрий. Этот бурый жир используется животным для теплопродукции и наиболее выражен у новорожденных или находящихся в зимней спячке млекопитающих, у которых он образует компактные массы, часто называемые гибернационными (зимовочными) железами, расположенными под кожей или в других специальных местах.
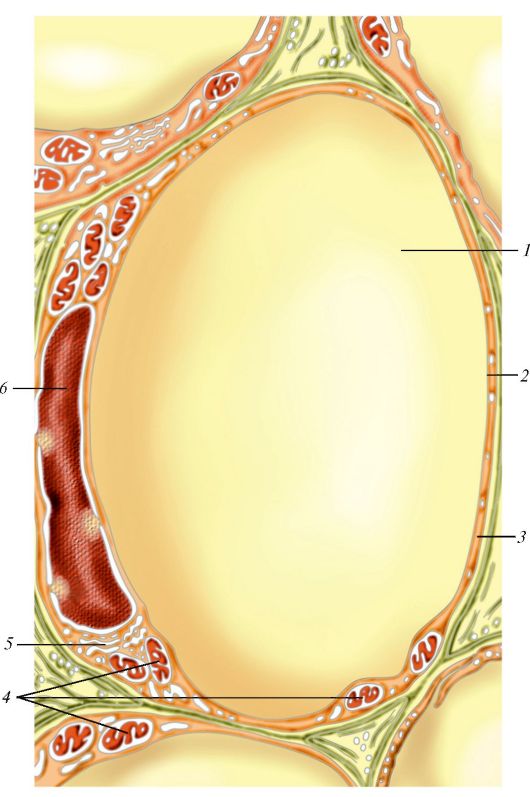
Строение однокапельного адипоцита: 1 — капля жира; 2 — цитоплазма адипоцита; 3 — базальная мембрана; 4 — митохондрии; 5 — сетчатый аппарат; 6 — ядро фибробласта.