Приветствую Вас Гость | RSS
Век млекопитающих - Age of Mammals
Меню сайтаФорма входа |
Строение и функции Агриотерии имели средние и крупные для медведей размеры: согласно расчетам, длина их тела могла достигать 2,7 м, высота в холке — 1,7 м, вес — 900 кг. Основная длина черепа составляет 40-48 см, длина m1 — 37-50,5 мм. Половой диморфизм в размерах хорошо выражен. Череп довольно низкий, с укороченным лицевым отделом. Профиль лба очень слабо вогнутый. Скуловые дуги очень мощные, широко расставленные. Глазницы широко расставлены. Сагиттальный гребень хорошо развит. Костное небо очень немного продолжается назад за зубы, широкое (ширина его у передних концов М1 значительно больше длины М1+М2), расширяющееся вперед: ширина у передних концов Р4 больше, чем у задних М2. Нижнечелюстная кость с глубокой премассетерной ямкой; в симфизном отделе имеется нижний выступ ("подбородок"); число подбородочных отверстий обычно увеличено. 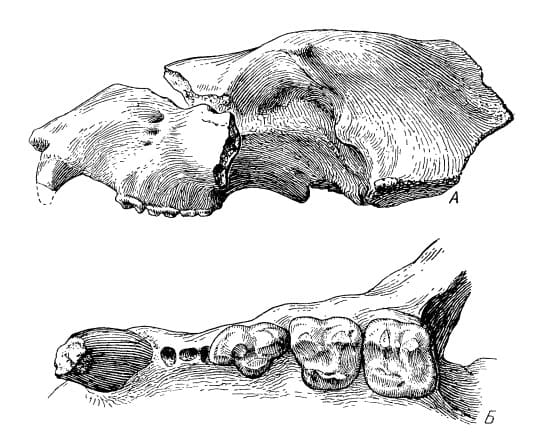 Прорисовки типового черепа агриотерия (Agriotherium sivalensis) из Сиваликов, Пакистан, и его левой верхнечелюстной кости с С, Р4-М2. Предкоренные с низкой коронкой, редуцированы в размере, однокорневые (иногда р3 с 2 корнями); вследствие этого длина докоренного ряда сильно уменьшена. Некоторые из предкоренных могут отсутствовать: обычно Р2/р2, реже Р1/p1 и РЗ/рЗ. Верхний хищнический зуб Р4 крупный, несколько длиннее M1, с сильным парастилем и высоким протоконом. Задний конец Р4 находится за подглазничным отверстием. Верхние коренные с более высокими коронками, чем у рода Indarctos. Длина M1 равна его ширине или привышает ее, язычный край зуба короче, чем щечной. М2 короткий, трапециевидных очертаний, назад не суживается, без развитого талона, так что уступает по длине M1; длина М2 немного меньше его ширины или равна ей и значительно меньше длины Р4+М1. р4 узкий (ширина его составляет около 1/2 длины), с 1 конусом (протоконидом) и простым талонидом. Нижний хищнический зуб m1 относительно короче, чем у Indarctos, его прегипокристид без дополнительного зубца и направлен вовнутрь, так что не сливается с щечным краем протоконида, как это имеет место у Indarctos. Форма язычных зубцов m1 варьирует. m2 с коротким талонидом, который упрощен по сравнению с таковым у Indarctos. Жевательная поверхность m3 округлой формы. m1 длиннее m2, m3 короче р4. 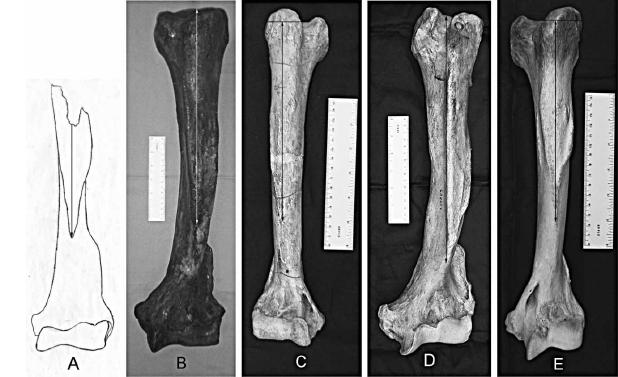 Плечевые кости: A — агриотерий (Agriotherium africanum), максимальный поперечный диаметр дистального конца 9,85 см; B — гигантский короткомордый медведь (Arctodus simus); C — гемицион (Hemicyon ursinus); D — прибрежный аляскинский бурый медведь (Ursus arctos gyas); E — тигр (Panthera tigris). Телосложение ариотериев более стройное, чем у современных медведей: их туловище менее бочкообразное, а конечности длиннее. Передняя часть туловища более развита, так что высота зверей в холке была гораздо больше высоты в крестце. Обе пары конечностей стопоходящие. Метаподиальные кости сходны с таковыми бурого медведя (Ursus arctos), однако ступни агриотерии судя по всему ставили прямо и не косолапили. В целом их морфология демонстрирует большое сходство с короткомордыми медведями рода Arctodus, что является следствием параллельной эволюции, так как эти медведи не состоят друг с другом в близком родстве и даже относятся к разным подсемействам. 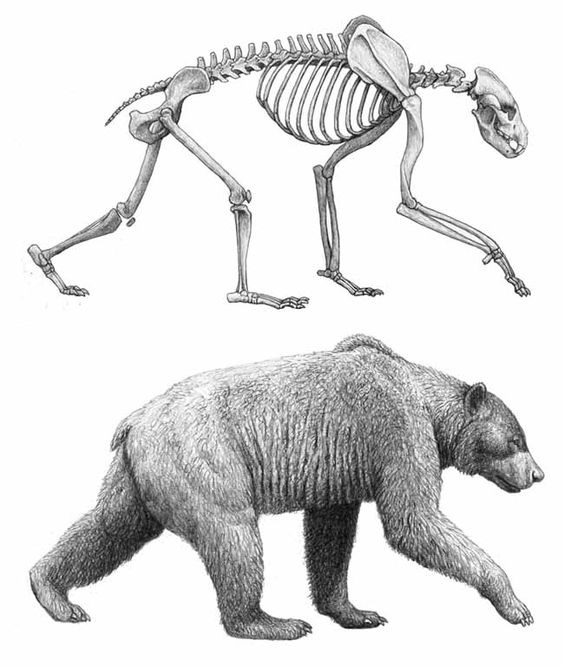 Прорисовка реконструированного скелета и реконструкция внешнего облика агриотерия (Agriotherium africanum). Автор — М. Антон. |
Поиск |