Аномодонты — Anomodontia
Класс †Синапсиды (Synapsida)
Подкласс †Терапсиды (Therapsida)
Надотряд †Динообразные (Dinomorpha)
Отряд †Аномодонты (Anomodontia)
Аномодонты (Anomodontia Owen, 1859) — отряд надотряда динообразных (Dinomorpha Ivakhnenko, 2003), происходящий из средней перми - ранней юры Африки, поздней перми - среднего или позднего триаса Мадагаскара, средней перми - позднего триаса Южной Америки, раннего - позднего триаса Антарктики, средней перми - позднего триаса Европы, средней перми - среднего триаса Азии, среднего - позднего триаса Северной Америки и, возможно, раннего триаса Австралии.
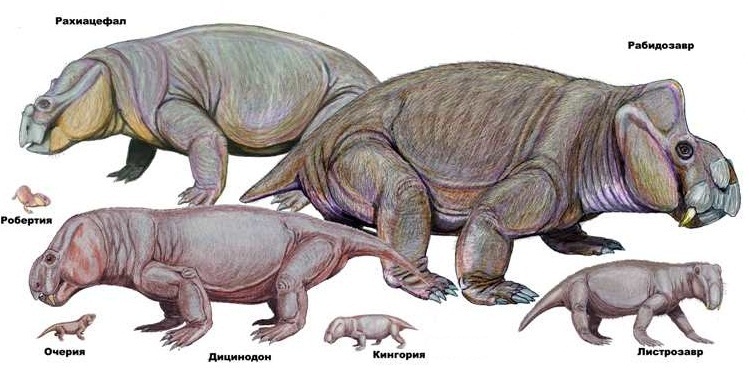
Разнообразие аномодонтов. Автор — Д. Богданов.
Название группы переводится с древнегреческого языка как «неправильно-» или «страннозубые».
Р. Оуэн (1859 г.) установил отряд аномодонтов, в котором объединил первых дицинодонтов, открытых незадолго перед этим, с ринхоцефалами. Избранное Р. Оуэном название располагало к включению в отряд самых разных рептилий с аберрантными типами озубления, но после Р. Брума (1905 г.) аномодонтами исследователи стали называть только дицинодонтов. Д. Ватсон и А. Ромер (1956 г.) пытались разделить всех терапсид на первичнохищных (Theriodontia) и первичнорастельноядных (Anomodontia), для чего присоединили к аномодонтам также и растительноядных диноцефалов — Tapinocephalia, но позднее А. Ромер (1966 г.) причислил к аномодонтам вообще всех диноцефалов. В понимании объема группы Д. Кинг (1988 г.) последовала за А. Ромером, но большинство ученых приняли аномодонтов в суженном объеме, присоединив к дицинодонтам также небольшие группы веньюковий и дромазавров (Хопсон и Бархьюзен (1986 г.), Кэрролл (1987 г.), Хопсон (1991 г.), Грин (1997 г.), Рыбчинская (2000 г.), Ивахненко (2004 г.)). Л.П. Татаринов последовал за последними авторами, рассматривая отдельно диноцефалов — и растительноядных, и хищных. Как единая группа диноцефалы рассматриваются и в монографии П.К. Чудинова (1983 г.).
Размеры от мелких до очень крупных: общая длина варьирует от 20 см до 4,5 м.
Как правило, череп короткий и крупный. Предглазничная часть черепа относительно укорочена, составляя не более 40 % от его общей длины, а также обычно несколько наклонена. Подобно диноцефалам, ноздри с конца морды сдвинуты назад и разделены сильно удлиненными восходящими отростками премаксилл, вклинивающимися между носовыми костями. Премаксиллы массивные и обычно сросшиеся друг с другом. Слезные кости умеренной длины, лобные достигают глазниц. Височное окно удлинено. Заглазничная и чешуйчатая кости смыкаются по верхнему краю височного окна. Скуловая дуга относительно грацильная, как бы вырезанная снизу. Она резко изгибается вверх, а затем круто опускается вниз у заднего конца. Внутренняя поверхность нижнего края скуловой дуги поворачивается кнаружи, так что наружные порции аддукторов оказываются открыты латерально и появляется возможность крепления мускулатуры к задне-наружному краю зубной кости. В результате конструкция приобретает черты, функционально аналогичные диапсидной схеме.
Субапсидная вырезка большая, в ее ограничении участвует отросток скуловой кости (т. н. «дифенестральная» височная область). Синдром связан с поднятием скуловой дуги вверх, так что в качестве височных окон функционируют верхняя — синапсидная — и нижнее — субапсидная вырезка. Чешуйчатая кость с длинным нижним отростком, прикрывающим снаружи квадратную кость и отделенным от костей лицевой части черепа глубокой вырезкой. Передний отросток чешуйчатой кости, входящий в скуловую дугу, очень длинный. Задний отросток скуловой дуги палочковидный и смещен вверх. Верхняя часть квадратно-скуловой кости расширена поперечно.
Теменные кости могут резко сужаться и образовывать сагиттальный гребень. Пинеальное отверстие всегда развито. Затылок широкий, обычно слегка наклонен верхней частью назад. Верхняя затылочная кость широкая. Околозатылочные отростки массивные, изогнутые на конце вниз. Овальное окно ушной капсулы смещено вентрально и соединено с внутренним ухом длинным перилимфатическим проходом. Челюстное сочленение вынесено далеко вниз и располагается примерно на уровне затылочного.
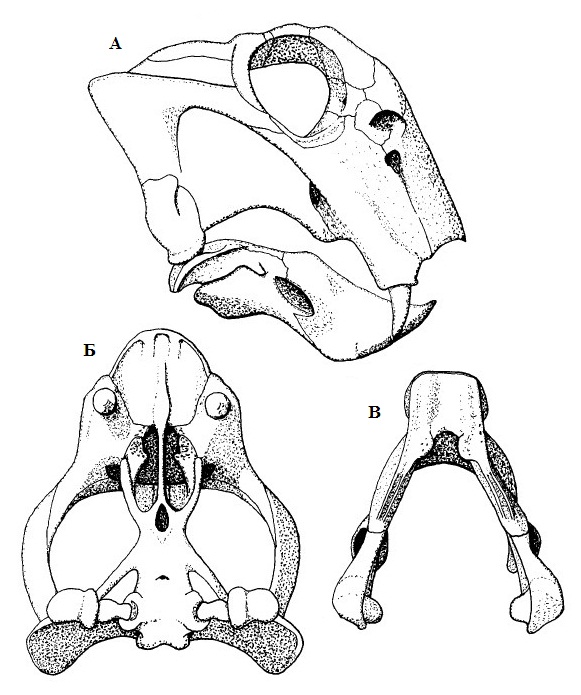
Прорисовки реконструированного черепа с нижней челюстью Lystrosaurus: А — сбоку, Б — снизу, В — нижняя челюсть сверху.
Имеется зачаточное вторичное небо, образованное разросшимися вентрально предчелюстньми и верхнечелюстными костями. Небные отростки премаксилл почти всегда развиты значительно и служили в качестве жесткой платформы при обработке пищи в ротовой полости. Небные кости разрастаются вперед по внутреннему краю верхнечелюстных и могут достигать предчелюстных, а также разрастаются назад от эктоптеригоидов. Вследствие развития вторичного неба хоаны смещены назад и разделены узкими, обычно срастающимися друг с другом желобчатыми сошниками. Базиптеригоидные отростки редуцированы. Крыловидные кости смыкаются непосредственно впереди и под телом базисфеноида и межптеригоидные ямы укорочены. Поперечные фланги крыловидных костей развиты слабо. Небные зубы отсутствуют.
Нижняя челюсть высокая, с выпуклым верхним краем наружным боковым мандибулярным окном. На наружной поверхности зубной кости (в задней ее половине) появляется площадка для прикрепления одного из нижнечелюстных аддукторов. Как и у диноцефалов, венечный отросток отсутствует. За исключением ряда базальных форм, артикулярная кость приобрела выпуклую сочленовную поверхность, что позволило нижней челюсти совершать пропалинальные (т. е. продольные) движения.
Челюстные зубы однообразные, обычно низкие и колышковидные, либо уплощенные сверху. Иногда зубы расположены несколькими рядами, зачастую их число сильно уменьшено. У продвинутых дицинодонтов образуется пара мощных верхнечелюстных клыков, а остальные челюстные зубы замещаются роговым клювом (признаки развития рогового покрова имеются, по крайней мере, на передней части нижней челюсти).
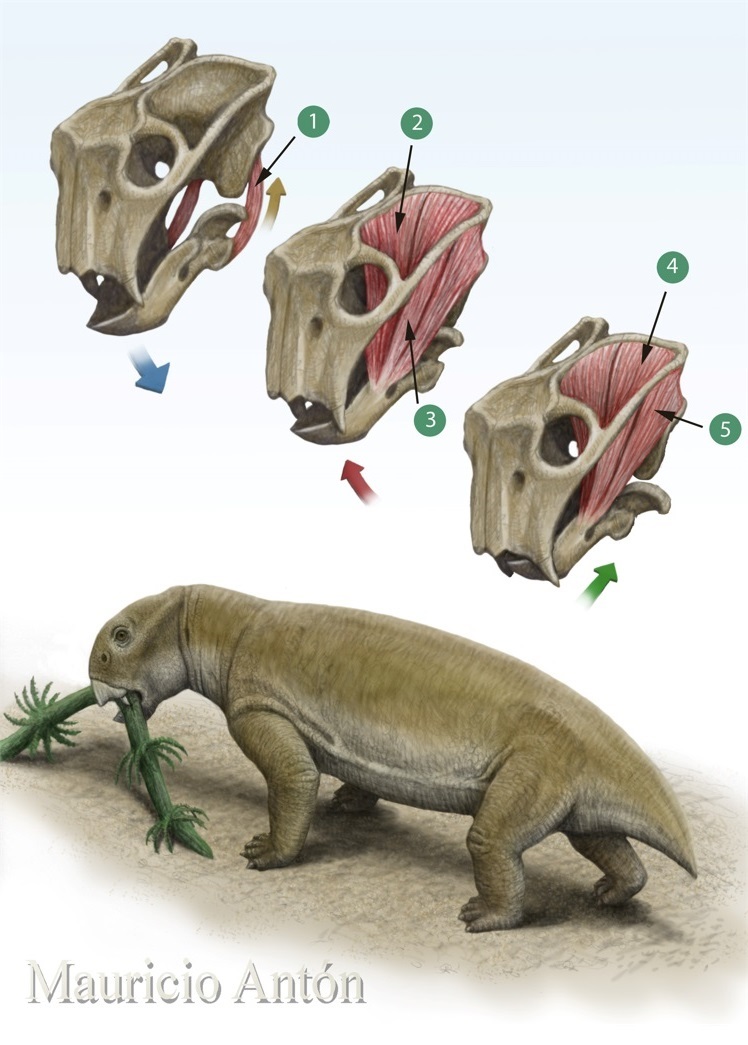
Реконструкция внешнего облика и схема действия челюстной мускулатуры Lystrosaurus. Сначала двубрюшная мышца (1) отводит вниз нижнюю челюсть. Пасть раскрывается, в нее помещается корм. Затем передние нижнечелюстные аддукторы (2, 3) смыкают челюсти, вызывая режущее движение и, как следствие, акт откусывания. И, наконец, в дело вступают задние аддукторы (4, 5), смещающие сомкнутые челюсти в горизонтальной плоскости, отчего откушенное разрывается, раздавливается, перетирается — проще говоря, пережевывается. Автор — М. Антон.
Посткраниальный скелет тяжело построенный. Позвонки глубоко амфицельные (вогнуты спереди и сзади). В плечевом поясе может сохраняться клейтрум. Коракоиды редуцированы или полностью утрачены. Конечности массивные. Как и у диноцефалов, внутренний трохантер бедренной кости не развит. Фаланговая формула маммального типа: 2 - 3 - 3 - 3 - 3. Иногда сохраняются гастралии.
Вероятно, первоначально аномодонты были насекомоядными, но потом перешли к настоящей растительноядности. Они первыми из терапсид начали не просто давить, а перетирать свою пищу во рту. Эффективная обработка грубой растительности, возможно, стала основным эволюционным приобретением данной группы и важным фактором ее диверсификации.
Обычно принято считать, что аномодонты связаны родством с растительноядными диноцефалами, что находит выражение во включении их некоторыми исследователями в один подотряд аномодонтов. Но у Т. Роу (1986 г.) на кладограмме аномодонты помещены между горгонопсами и тероцефалами, и он основывается, в частности, на разрастании и у териодонтов, и у аномодонтов височной ямы назад и наружу. Дж. Хопсон (1991 г.) утверждает все же, что аргументов, приводимых Т. Роу, недостаточно, чтобы отказаться от диноцелафовой гипотезы, которую считает более вероятной. Отметим все же и мнение Р. Кэрролла (1987 г.), по словам которого оснований для объединения аномодонтов с любой группой примитивных терапсид пока не имеется.
Ниже представлено несколько схем филогении аномодонтов.
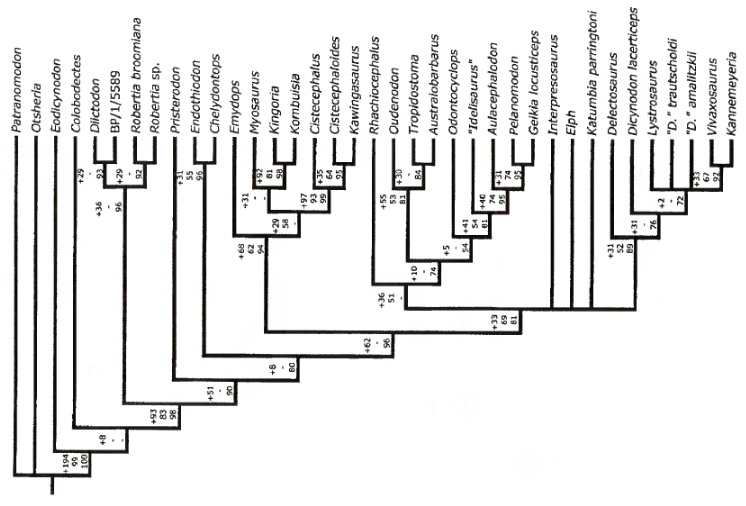
Схема филогении аномодонтов (по Ангельчику, 2007).

Схема филогении аномодонтов (по Фребишу, 2007).
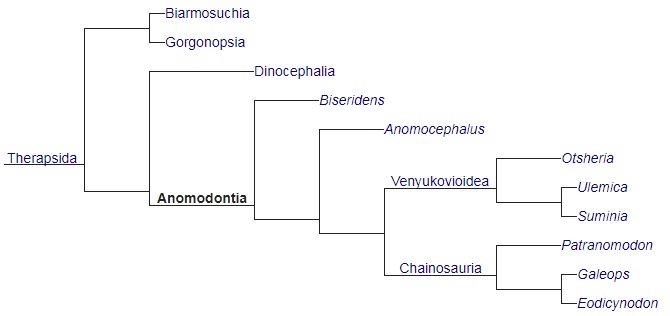
Кладограмма, иллюстрирующая родственные связи аномодонтов (по Лю и др., 2009)

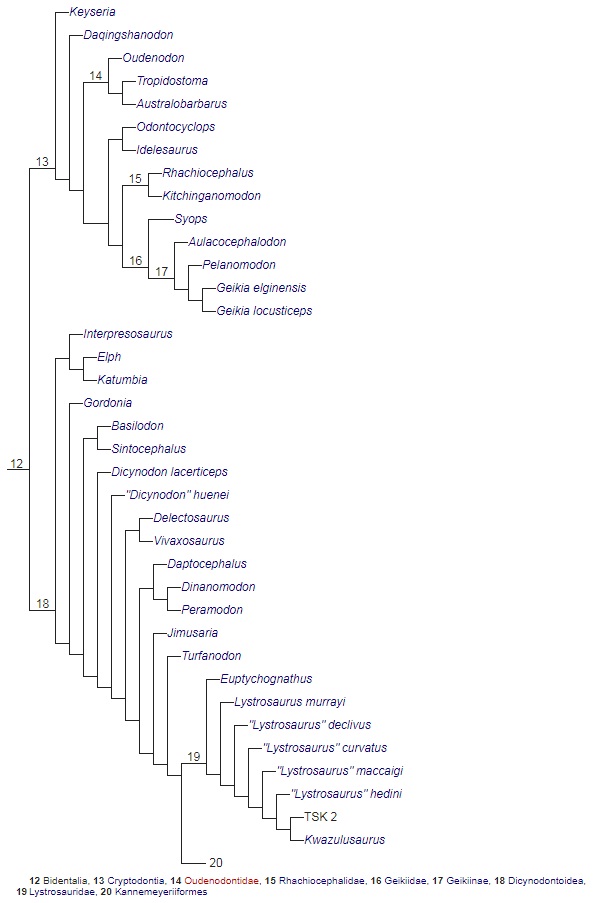
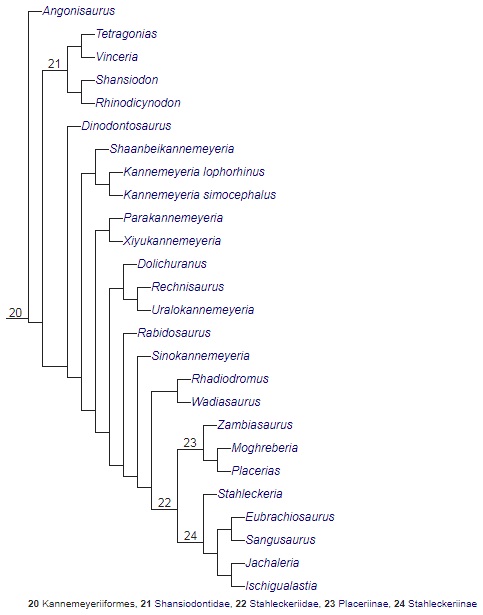
Кладограмма, иллюстрирующая родственные связи аномодонтов (по Каммереру и др., 2013).
Ссылки
Аномодонты (Википедия) перейти
Anomodontia (Paleobiology Database) перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Иванов А. О., Черепанов Г. О. Ископаемые высшие позвоночные. — СПб.: Издательство Санкт-Петербургского университета, 2001
Ивахненко М. Ф. Тетраподы Восточно-Европейского плаката — позднепалеозойского территориально-природного комплекса. — Пермь, 2001
Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1 / Отв. ред. Ивахненко М. Ф. и Курочкин Е. Н. — М.: ГЕОС, 2008
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГЕОС, 2009