Археоиндри — Archaeoindris
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Суперотряд Эвархонтоглиресы (Euarchontoglires)
Грандотряд Эвархонты (Euarchonta)
Миротряд Приматоподобные (Primatomorpha)
Отряд Приматы (Primates)
Подотряд Мокроносые приматы (Strepsirrhini)
Инфраотряд Лемурообразные (Lemuriformes)
Надсемейство Лемуроиды (Lemurоidеа)
Семейство †Палеопропитековые (Palaeopropithecidae)
Род †Археоиндри (Archaeoindris)
Вид †Archaeoindris fontoynontii
Археоиндри (Archaeoindris Standing, 1909) — монотипический род семейства палеопропитековых (Palaeopropithecidae Tattersall, 1973), происходящий из голоцена Мадагаскара.

Реконструкция внешнего облика археоиндри (Archaeoindris fontoynontii). Автор — Г. Угуэто.
Синоним: Lemuridotherium Standing, 1910.
Единственный установленный вид — Archaeoindris fontoynontii Standing, 1909 (тип). Его остатки обнаружены в центральной части Мадагаскара. Они датируются голоценом. Синонимы: Archaeoindris fontoynonti Standing, 1909; Lemuridotherium madagascariensis Standing, 1910.
История изучения
Рассматриваемый таксон был установлен британским палеонтологом Г. Стэндингом в 1909 г. В основу описания легли 3 образца: нижняя челюсть с полным набором зубов и 2 фрагмента верхней челюсти — левый и правый. Стэндинг причислил нового лемура к подсемейству "Indrisinées" и дал ему родовое имя Archaeoindris, образованное от древнегреческого корня ἀρχαῖος ("древний") и названия "индри" — в силу предполагаемого близкого родства с этими современными лемурами. Видовое имя fontoynontii присвоено в честь французского врача, этнолога и историка А.- М. Фонтуанона — президента Малагасийской академии наук, руководившего раскопками, в ходе которых и был обнаружен археоиндри.
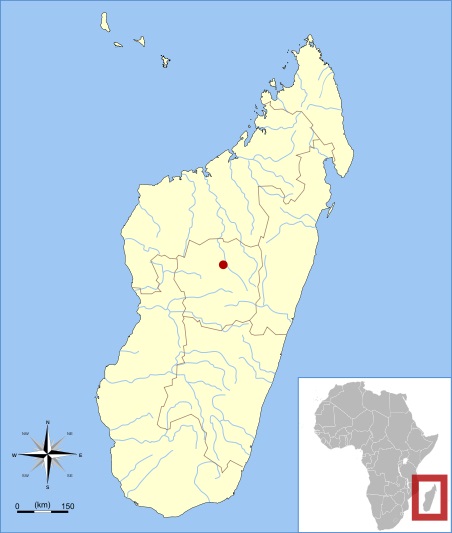
Карта местонахождений остатков археоиндри (Archaeoindris fontoynontii).
Типовые образцы были найдены в пещере Ампасамбазимба на Центральном нагорье. Там же впоследствии были сделаны все остальные находки, ассоциируемые с данным видом. (В середине 2010-х гг. сообщалось об открытии в национальном парке Циманампецуца подводного месторождения, содержащего, по-видимому, в числе прочих и кости археоиндри.) Количество находок остается очень небольшим, главнейшей из них стало обнаружение французским палеонтологом Ш. Ламбертоном в первой половине 1930-х гг. полного черепа с очевидно принадлежащей ему нижней челюстью.
При этом в публикации Ламбертона за 1934 г. были сделаны ключевые ошибки: обнаруженные им посткраниальные останки, приписанные археоиндри, в дальнейшем были определены как кости мегаладаписа — довольно далекого в эволюционном плане субфоссильного лемура (возможно, что причиной ошибки стала принадлежность этих костей более крупному, чем известные к тому времени, экземпляру мегаладаписа). На основании ошибочного определения Ламбертон описал строение тела и предполагаемый образ жизни археоиндри как близкие к таковым у мегаладаписов; эта ошибка позже неоднократно воспроизводилась в разных научных работах. Напротив, по бедренной кости, впоследствии опознанной как принадлежащая археоиндри, был первоначально описан отдельный род — Lemuridotherium, позднее признанный младшим синонимом Archaeoindris.
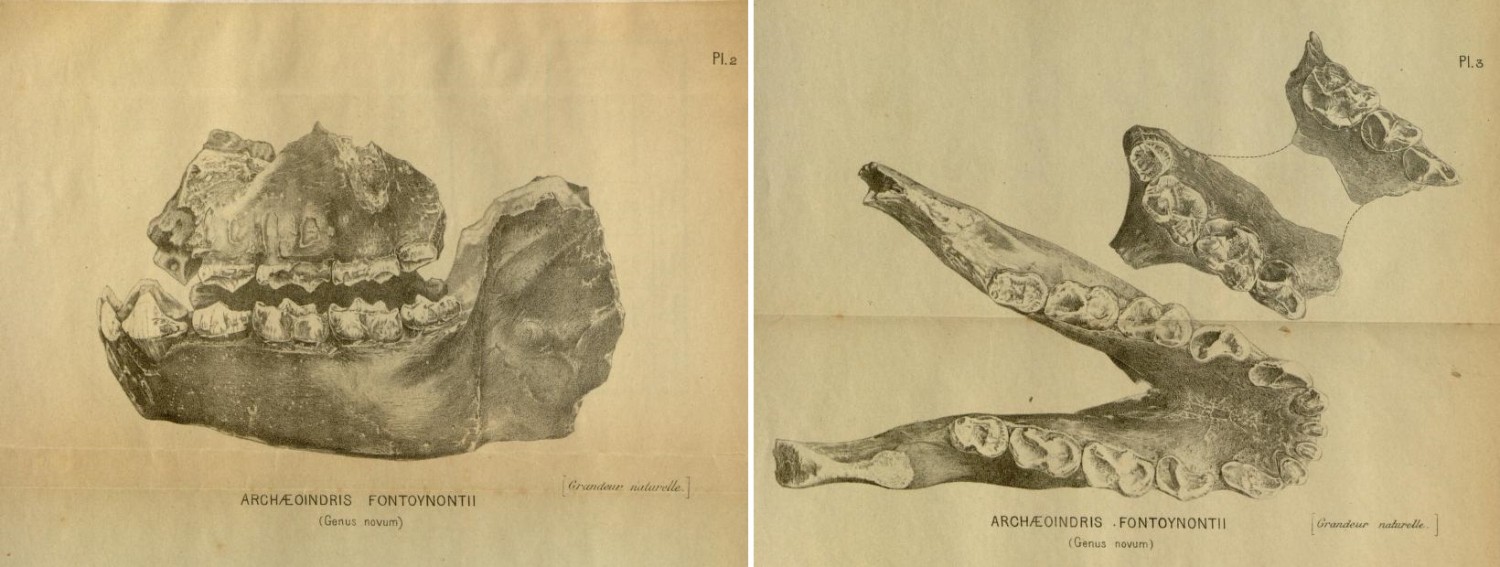
Прорисовки типового черепного материала археоиндри (Archaeoindris fontoynontii).
Ошибочная идентификация отдельных костей последовательно опровергалась различными исследователями между 1936 и 1977 гг. В обзорной статье 1988 г. сообщалось, что, помимо черепа и отдельных челюстей, с уверенностью можно утверждать о принадлежности археоиндри всего 6 костей (целых и обломков) посткраниального скелета, причем 4 из них принадлежат неполовозрелой особи — по всей видимости, только одной. Ограниченное количество известных остатков археоиндри делает его наименее изученным из всех субфоссильных лемуров. В настоящее время этот род относят к семейству палеопропитековых (Palaeopropithecidae) и включают в него единственный вид Archaeoindris fontoynontii. Внутри этого семейства археоиндри был, по всей видимости, ближе всего к следующему по размеру роду — палеопропитекам (Palaeopropithecus), о чем свидетельствует ряд общих черт в строении черепа и устройстве зубов.
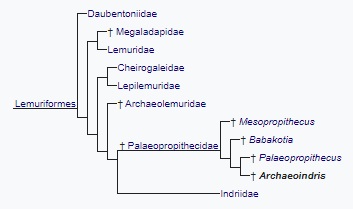
Место археоиндри (Archaeoindris fontoynontii) в систематике лемурообразных.
Строение и функции
Археоиндри является крупнейшим известным науке мокроносым приматом: его череп достигает 27 см в длину, а общая длина тела оценивается в 1,5 м. Вес предположительно мог составлять 150-200 кг, что сопоставимо с массой тела крупных самцов горилл (Gorilla).


Череп с нижней челюстью археоиндри (Archaeoindris fontoynontii).
Череп по габаритам короче, но шире, чем черепа мегаладаписов (Megaladapis), и массивней и глубже, чем черепа палеопропитеков (Palaeopropithecus). Как и для черепов сифак (Propithecus), характерна меньшая (относительно общих размеров) длина и большая глубина; возможно, что подобное вытягивание черепа в высоту было связано с необходимостью компенсировать высокие нагрузки при пережевывании пищи.
Лицевой отдел черепа несколько вытянут и сужен, мозговой небольшой. Наблюдается ряд черт, общий с палеопропитеками: верхние части предчелюстных костей вместе с носовыми костями образуют парные выступы над носовым отверстием, глазницы посажены довольно близко, имеется выраженное сужение черепа в заглазничной области, твердое небо прямоугольное, наружный слуховой проход трубчатый, слуховой барабан уплощен. В то же время по целому ряду параметров череп археоиндри ближе к гипотетическим общим предкам палеопропитековых и индриевых и менее специализирован, чем у палеопропитеков. Лицевой отдел относительно короче. Слабее выражено утолщение костей вокруг глазниц, сами глазницы не так сдвинуты к затылку, и вверх. Черепной свод маленький относительно общих размеров; меньший объем черепной коробки по сравнению с обезьянами сопоставимой величины в целом характерен для субфоссильных лемуров. Имеется широкий сагиттальный гребень, а также затылочный гребень. Строение среднего уха примитивно — например, присутствует костяной канал, соединяющий барабанную полость с внешней средой. Длинный нижнечелюстной симфиз срастался рано.
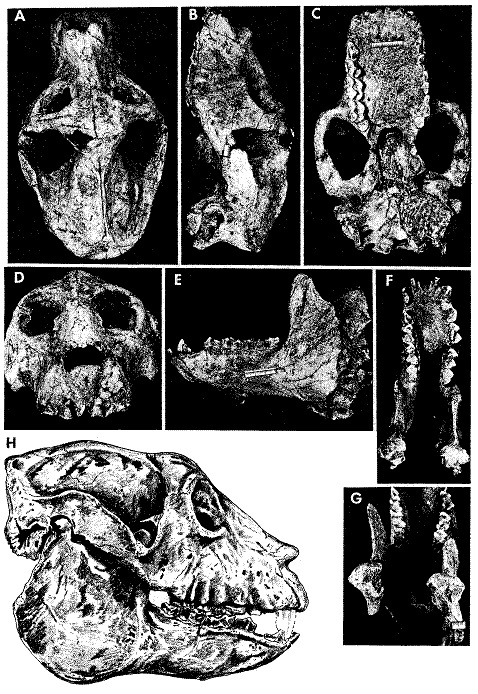
Череп с нижней челюстью археоиндри (Archaeoindris fontoynontii) и его прорисовка.
Зубная формула — I2/i1, C1/c1, P2/р2, M3/m3 — сходна с таковой у других палеопропитековых (Palaeopropithecidae) и индриевых (Indriidae). Так же, как у палеопропитеков, и в отличие от менее крупных бабакотии (Babakotia) и мезопропитеков (Mesopropithecus) (и всех современных мокроносых приматов, кроме руконожки (Daubentonia madagascariensis)) нижние резцы не образуют "зубной гребенки" — эти зубы у данных 2 родов короткие и тупые. Нижние предкоренные разделены небольшой диастемой. Как и у палеопропитеков, передние коренные уплощены в щечно-язычном направлении, М3 редуцирован, эмаль щечных зубов мелкогородчатая. Эти дентальные характеристики археоиндри разделяет не только с палеопропитеками, но и с некоторыми другими представителями своего семейства, а также сифаками. Коронки зубов более высокие, а эмаль несколько менее морщинистая, чем у палеопропитеков.
Единственной известной целой костью посткраниального скелета, принадлежавшей взрослой особи археоиндри, является бедренная кость. Взрослой особи принадлежит также поврежденная плечевая кость; помимо этого, известны 4 длинные кости конечностей подростковой особи.

Этапы портретной реконструкции археоиндри (Archaeoindris fontoynontii). Автор — В. Симеоновски.
Плечевую кость археоиндри отличают от костей мегаладаписов слабовыраженные дельтовидная бугристость и узел крепления плечелучевой мышцы, мелкая межбугорковая борозда, а также прямые, а не вогнутые линии медиального края. Диафиз почти круглый в сечении (у мегаладаписов треугольный). Локтевая кость, известная только для молодой особи, вероятно, сопоставима по длине с плечевой, полуцилиндрическая в сечении диафиза (у мегаладаписов Т-образная), менее массивная. Широкая блоковидная вырезка напоминает устройство локтевой кости у палеопропитеков.
Бедренная кость взрослого археоиндри относительно короткая, но чрезвычайно массивная, с огромной головкой, на которой отсутствует впадина. Большой вертел редуцирован (как у палеопропитеков). Шеечно-диафизарный угол (угол пересечения диафиза и шейки бедренной кости) высокий (164 ° — у палеопропитеков в среднем порядка 168 °). Диафиз в сечении овальный (сильнее сплюснутый у кости взрослой особи, чем у подростковой). Рядом с малым вертелом отсутствует треугольный выступ, заметный на бедренных костях мегаладаписов (но так же точно отсутствующий у палеопропитеков). III-й вертел практически отсутствует, на его месте едва заметная складка.

Бедренная кость археоиндри (Archaeoindris fontoynontii).
Интермембральный индекс (соотношение длин плечевой и бедренной костей) установить точно не представляется возможным ввиду отсутствия целой плечевой кости взрослой особи. Это соотношение несомненно превышает 100 — плечевая кость длиннее бедренной, как и локтевая кость у подростковой особи, хотя разница в длине передней и задней конечностей, по-видимому, не столь велика, как у палеопропитеков (интермембральный индекс последних составляет 138-147, являясь наибольшим у приматов).
Образ жизни
Открыватель археоиндри Стэндинг предполагал, что этот примат вел, несмотря на свои размеры, древесный образ жизни, подобный образу жизни палеопропитеков, то есть медленно перелезал с ветки на ветку, много времени проводя в висячем положении. Ошибочное приписывание археоиндри некоторых костей палеопропитеков послужило подтверждением этой модели поведения и для Ламбертона, а также более поздних авторов, опиравшихся на его работу 1934 г. Вместе с тем Ламбертон, определив, что часть известных костей сближает этот род с палеопропитеками, одновременно высказал предположение, что его представители могли вести наземный образ жизни, наподобие многих вымерших ленивцев. Эту модель в 1980 г. поддержал американский антрополог У. Юнгерс, указав, что уже в силу значительных габаритов археоиндри не мог висеть спиной вниз и вынужден был проводить много времени на земле.

Реконструкция внешнего облика археоиндри (Archaeoindris fontoynontii). Автор — С. Нэш.
При этом на способность лазания по деревьям указывают относительно большая свобода движения тазобедренного сустава (о которой свидетельствует большой шеечно-диафизарный угол бедренной кости) и значительная длина плечевой и локтевой костей по сравнению с бедренной костью. В отсутствие находок костей кисти и стопы определить степень специализации конечностей археоиндри для лазания и висения (у палеопропитеков очень высокую) пока не представляется возможным. Поэтому в более поздних публикациях этот лемур рассматривается как, вероятно, адаптированный к лазанию по древесным стволам и крепким ветвям, но проводящий достаточно много времени на земле, как в поисках пищи, так и с целью перемещения от одного дерева к другому; при этом передвижение по земной поверхности, скорее всего, было медлительным и даже неуклюжим.


Археоиндри (Archaeoindris fontoynontii) и серый лемур (Hapalemur grisues). Автор — Р. Схоутен.
Характер микроизноса зубов, установленный по известным остаткам археоиндри (редкие тонкие царапины, низкая концентрация ямковидных щербин), указывает на то, что это был высокоспециализированный листоядный примат, в рационе которого семена присутствовали, но не составляли существенной части. Из современных приматов схожая диета встречается у некоторых тонкотелых обезьян (в том числе очкового тонкотела (Trachypithecus obscurus), гривистого тонкотела (Trachypithecus cristatus) и королевского колобуса (Colobus polykomos)), а также у части крупных современных лемуров (диадемового сифаки (Propithecus diadema)), а из ископаемых видов — не у палеопропитеков, а у более мелкого вида того же семейства, Mesopropithecus pithecoides.

Археоиндри (Archaeoindris fontoynontii) в естественной среде обитания. Автор — Р. Евсеев.
Небольшие относительно размеров тела глазницы, сравнимые по площади с глазницами горилл, указывают на дневную активность археоиндри. При этом соотношение ширины канала зрительного нерва к размерам глазницы (т. н. индекс зрительного отверстия) у археоиндри и других крупных субфоссильных лемуров — мегаладаписов (Megaladapis) и гадропитека (Hadropithecus) — было очень низким по сравнению с обезьянами (и даже по сравнению с более мелкими родственниками из числа палеопропитековых), что свидетельствует о меньшей резкости зрения.


Самка археоиндри (Archaeoindris fontoynontii) с детенышем.
Вымирание
Вымирание
Конкретные временные рамки существования археоиндри не определены. Стратиграфическая датировка отложений в пещере Ампасамбазимба варьирует от 9 до 5,3 тыс. лет. При этом, согласно радиоуглеродным датировкам, часть найденных остатков археоиндри датируется возрастом 2-2,5 тыс. лет, а другие — 8 тыс. лет назад, что делает вероятным его исчезновение еще до заселения Мадагаскара человека порядка 1,5 тыс. лет назад. Если пересечение все же произошло, человеческая деятельность — как охота, так и сведение лесов под сельскохозяйственные нужды — должна была быстро привести к исчезновению этого и так достаточно редкого, медленно размножающегося и чувствительного к переменам окружающей среды лемура.
Изменение среды обитания, как показывают исследования в центральной части Мадагаскара, в значительной степени являлось результатом климатических изменений, приведших к исчезновению влажных лесов и замене их безлесной саванной уже к моменту прихода человека. Однако костные остатки, в том числе из месторождения Ампасамбазимба, доказывают, что сами по себе климатические изменения не привели к полному исчезновению мадагаскарской мегафауны; таким образом, по-видимому, оно стало результатом взаимодействия природных факторов и человеческой деятельности.
Ссылки
Археоиндри (Википедия) перейти
Archaeoindris (Paleobiology Database) перейти
Литература
Дробышевский С. Достающее звено. В 2 т. Т. 1. Обезьяны и все-все-все. — Corpus, 2017
Каландадзе Н. Н., Шаповалов А. В. Судьба мегафауны наземных экосистем Мадагаскарской зоогеографической области в позднем антропогене.
Мадагаскар / Науч. ред. Б. Д. Васильев. — М.: Прогресс, 1990
Body Size in Mammalian Paleobiology: Estimation and Biological Implications / Ed. by Damuth J., MacFadden B. J. — Cambridge University Press, 1990
Cenozoic mammals of Africa / Ed. by Werdelin L., Sanders W. J. — University of California Press, 2010
Crowley B. E. A refined chronology of prehistoric Madagascar and the demise of the megafauna. Quaternary Science Reviews 29, 2010
Dunkel A. R., Zijlstra J. S., Groves C. P. Giant rabbits, marmosets, and British comedies: etymology of lemur names, p. 1. Lemur News 6, 2011/2012
Elwyn Simons: A search for origins / Ed. by Fleagle J. G., Gilbert C. C. — New York: Springer, 2008
Encyclopedia of Human Evolution and Prehistory / Ed. by Delson Е., Tattersall I., Van Couvering J., Brooks A. S. — Taylor & Francis, 2000
Godfrey L. R., Jungers W. L. The Extinct Sloth Lemurs of Madagascar. Evolutionary Anthropology 12, 2003
Godfrey L. R., Semprebon G. M., Jungers W. L., Sutherland M. R., Simons E. L., Solounias N. Dental use wear in extinct lemurs: evidence of diet and niche differentiation. Journal of Human Evolution 47 (3), 2004
Godfrey L. R., Schwartz G. T., Samonds K. E., Jungers W. L., Catlett K. K. The Secrets of Lemur Teeth. Evolutionary Anthropology Issues News and Reviews 15 (4), 2006
Goodman M. S., Jungers W. L. Extinct Madagascar: Picturing the Island's Past. — The University of Chicago Press, 2014
Jungers W. L., Godfrey L. R., Simons E. L., Chatrath P. S. Phalangeal curvature and positional behavior in extinct sloth lemurs (Primates, Palaeopropithecidae). Proceedings of the National Academy of Sciences of the United States of America 94 (22), 1997
Karanth K. P., Delefosse T., Rakotosamimanana B., Parsons T. J., Yoder A. D. Ancient DNA from giant extinct lemurs confirms single origin of Malagasy primates. Proceedings of the National Academy of Sciences of the United States of America 102 (14), 2005
Lemurs: Ecology and Adaptation / Ed. by Gould L., Sauther M. L. — Springer Science & Business Media, 2006
Mittermeier R. A., Louis E. E., Richardson M., Schwitzer C., Langrand O., Rylands A. B., Hawkins F., Rajaobelina S., Ratsimbazafy J., Rasoloarison R., Roos C., Kappeler P. M., MacKinnon J. Lemurs of Madagascar (3rd ed.). — Conservation International, 2010
Nowak R. M. Walker's Primates of the World. — Baltimore, MD: The Johns Hopkins University Press, 1999
Primate evolution and human origins / Ed. by Ciochon R. L, Fleagle J. G. — Hawthorne, NY: Aldine de Gruyter, 1987
Primate evolution and human origins / Ed. by Ciochon R. L, Fleagle J. G. — Routledge, 2017
Reconstructing Behavior in the Primate Fossil Record / Ed. by Plavcan J. M., Kay R. F., Jungers W. L., van Schaik C. P. — Springer Science & Business Media, 2012
Standing H. F. Subfossiles provenant des fouilles d’Ampasambazimba. Bulletin de l’Academie Malgache 6, 1909
Szalay F., Delson E. Evolutionary history of the Primates. — New York: Academic Press, 1979
Tattersall I. The Primates of Madagascar. — New York: Columbia University Press, 1982
The Natural History of Madagascar / Ed. by Goodman S. M., Benstead J. P. — University of Chicago Press, 2003
The Primate Fossil Record / Ed. by Hartwig W. C. — Cambridge: Cambridge University Press, 2002
Vuillaume-Randriamanantena M. The taxonomic attributions of giant subfossil lemur bones from Ampasambazimba: Archaeoindris and Lemuridotherium. Journal of Human Evolution 17 (4), 1988