Археолемуры — Archaeolemur
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Суперотряд Эвархонтоглиресы (Euarchontoglires)
Грандотряд Эвархонты (Euarchonta)
Миротряд Приматообразные (Primatomorpha)
Отряд Приматы (Primates)
Подотряд Мокроносые приматы (Strepsirrhini)
Инфраотряд Лемурообразные (Lemuriformes)
Надсемейство Лемуроиды (Lemurоidеа)
Семейство †Археолемуровые (Archaeolemuridae)
Род †Археолемуры (Archaeolemur)
Вид †Archaeolemur edwardsi
Вид †Archaeolemur majori
Археолемуры (Archaeolemur Filhol, 1895) — род семейства археолемуровых (Archaeolemuridae Grandidier, 1905), происходящий из голоцена Мадагаскара. Установлены 2 вида.

Реконструкция внешнего облика археолемура (Archaeolemur majori). Автор — Smokeybjb.
Синонимы: Bradylemur Grandidier, 1899; Globilemur Forsyth Major, 1898; Nesopithecus Forsyth Major, 1896.
Строение и функции
Размеры археолемуров достаточно крупные: их длина их черепа составляет 13-15 см, а вес оценивается в 15-25 кг.
Археолемуры являлись менее специализированным представителями семейства археолемуровых (Archaeolemuridae), чем гадропитеки (Hadropithecus). По строению черепа они походят на современных индриевых (Indriidae). Сходство между черепом вида Archaeolemur majori и черепами индриевых настолько явно, что даже предлагалось выделить этот вид в отдельный род Protoindri. В то же время, наблюдаются специфические особенности в строении зубочелюстного аппарата археолемуров. Их нижнечелюстной симфиз вертикальный и сросшийся, в отличии от пологого несросшегося симфиза индриевых.
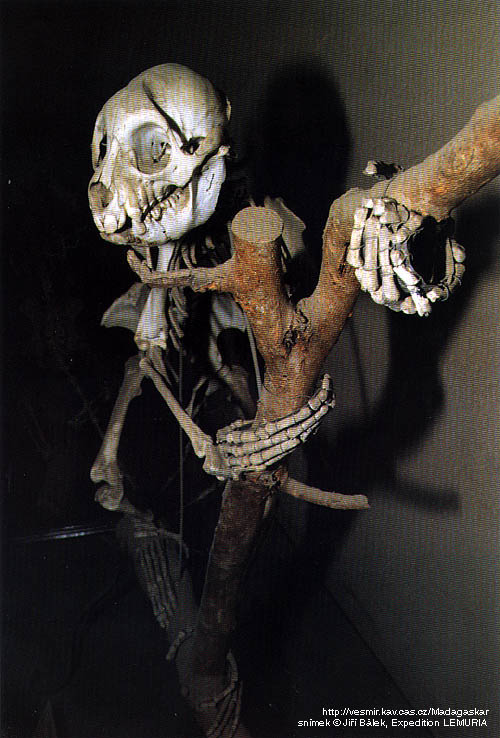

Скелет археолемура (Archaeolemur majori).
Зубная формула: I2/i1, C1/c1, P3/р3, M3/m3. Центральные верхние резцы сильно увеличены, расширены и, вероятно, не были прикрыты губой при жизни. Нижние резцы также крупные. Хорошо развиты клыки, особенно нижние. Предкоренные сжаты с боков и формируют как бы продольное лезвие, удобное для срезания растительности. Коренные низкие и широкие, в каждом углу с острым выступом, которые соединяются 2 гребешками (1 с внешней, 1 с внутренней стороны). Таким образом, у археолемуров наблюдается билофодонтное строение зубов, отмеченное помимо него лишь у обезьян Старого Света. Подобная адаптация, способствующая качественному измельчению корма, может свидетельствовать о широком спектре потребляемой пищи — в частности, жесткой растительности, о преобладании которой в рационе свидетельствует высокая степень износа коренных. Многочисленные находки позволили установить, что очередность прорезывания постоянных зубов весьма близка к таковой индриевых.
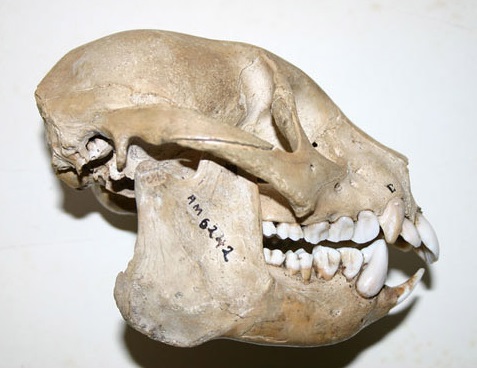
Череп археолемура (Archaeolemur majori).
Мощное туловище поддерживали относительно короткие для лемуров конечности (они короче, чем у короткохвосто индри (Indri indri), и это учитывая то, что последний по меньшей мере вдвое мельче археолемуров). Задние ноги несколько длиннее передних (интермембральный индекс — 92). Хватательная способность кисти и стопы снижена: по степени искривленности фаланги пальцев уступают таковым как вымерших, так и современных лемуров и скорее напоминают фаланги павианов (Papio) и горилл (Gorilla), ведущих преимущественно наземный образ жизни. Половой диморфизм не выражен.
Обильные остатки археолемуров позволили выделить фрагменты их ДНК и провести достаточно полные исследования генетического материала.
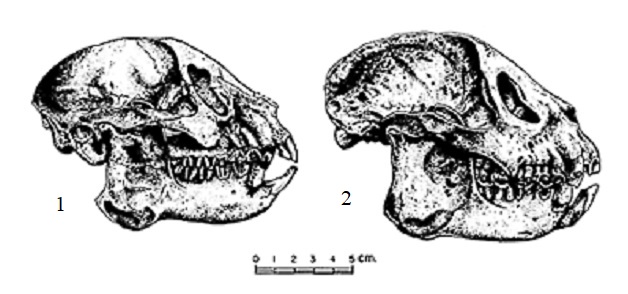
Прорисовки черепов археолемуров (Archaeolemur): 1 — Archaeolemur majori; 2 — Archaeolemur edwardsi.
Состав
Archaeolemur edwardsi Filhol, 1895 — вид, известный из голоцена центральных районов Мадагаскара; возможно (принадлежность остатков не подтверждена окончательно), также жил на севере, западе и юго-западе острова. Остатки происходят из местонахождений Ампасамбазимба, Ампоза-Анказоабо, Масинандраина, Бело-сур-мер, Морарано-Бетафо, Самбаина, Вакинанкаратра (Центральный Мадагаскар). Вероятные остатки обнаружены в местонахождениях Амбулисатра, Ампарихингидро, Андрахомана, Анджохибе, Анзухикели, Анкарана, Бунго-Тсиманиндроа, Мт. дес Франкайс.
Их двух видов отличается более крупными размерами и массивным сложением. Череп имеет около 15 см в длину, максимальный вес оценивается в 25 кг. Череп имеет хорошо развитый сагиттальный гребень и массивный затылочный бугор. Индексы пропорций конечностей: интермембральный — 92, гумерофеморальный — 82-85, брахиальный — 107, круральный — 86.
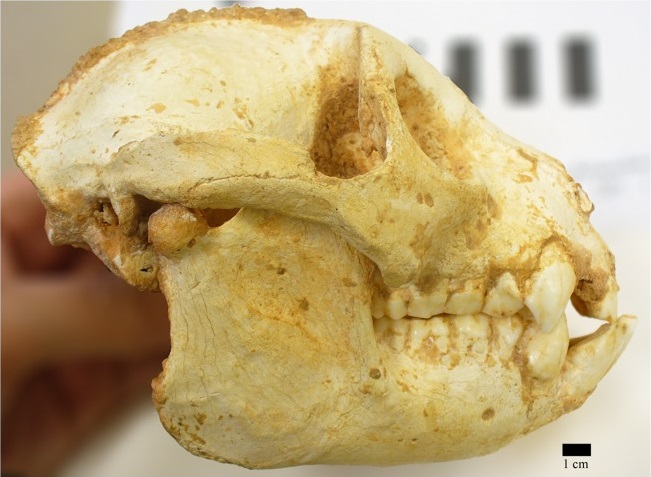
Череп археолемура (Archaeolemur edwardsi).
Archaeolemur majori Filhol, 1895 — вид, известный из голоцена южной и западной части Мадагаскара; возможно также, что он обитал в центре и на севере острова. Типовое местонахождение — Бело-сур-мер на западе Мадагаскара.
Более мелкий и изящно сложенный вид. Длина черепа составляет около 13 см, вес оценивается примерно в 15 кг. Череп с менее развитым рельефом: сагиттальный и затылочный гребни практически не выражены. Индексы пропорций конечностей: интермембральный — 92, гумерофеморальный — 85-86, брахиальный — 100, круральный индекс — 94.

Черепа археолемура (Archaeolemur majori).
В последние десятилетия XX в. на севере и северо-западе Мадагаскара были обнаружены новые остатки, определенные лишь до рода и предварительно обозначенные Archaeolemur spр. По ряду признаков эти находки несколько отличаются не только от известных видов, но и между собой. Поэтому вполне вероятно, что в будущем будет выделен еще 1 или даже 2 вида археолемуров.
Возраст и распространение
Абсолютный возраст костных остатков археолемуров варьирует в диапазоне от 2850 до 1020 ± 50 лет. Датировка, полученная по окаменелым испражнениям, ассоциированным с остатками ювенильной особи из местонахождения Анзухикели на севере Мадагаскара, составила 830 ± 60 лет.
Полуназемный образ жизни, высокая мобильность и широкие вкусовые предпочтения археолемуров способствовали значительному распространению этих приматов — их остатки известны из более чем 20 местонахождений, разбросанных почти по всему Мадагаскару. Археолемуры изучены гораздо лучше, чем другой известный представитель их семейства — гадропитек (Hadropithecus stenognathus).
Образ жизни
Археолемуры представляли собой четвероногих полуназемных животных, по телосложению и образу жизни несколько напоминавших павианов (Papio). Ничто в их анатомии не указывает на способность к быстрому бегу. В отличие от гадропитека и даже от современных лемуров, ведущих дневной образ жизни, археолемуры, по-видимому, обладали более острым зрением (их расчетный зрительный индекс превышает таковой у сифак (Propithecus) и даже шимпанзе (Pan)). Обитали археолемуры главным образом в лесистых районах.

Археолемур (Archaeolemur edwardsi) в естественном окружении. Автор — Smokeybjb.
В рацион археолемуров прежде всего входили фрукты, листья, цветы, стебли и другие мягкие части растений. В то же время, характерные следы износа на многих из найденных зубов свидетельствуют о длительном питании жестким растительным материалом, который могли составлять семена, плоды с твердым околоплодником и, возможно, кора деревьев. Анализ копролитов позволил установить, что помимо растительной пищи также поедались улитки, членистоногие и небольшие позвоночные животные (например, летучие мыши и мелкие лемуры).
Вымирание
Археолемуры были одними из последних выживших субфоссильных лемуров в фауне Мадагаскара. Логично предположить, что вследствие довольно крупных размеров они являлись желанной добычей для малайских переселенцев. Выжигание лесов под поля и пастбища влекла за собой сокращение естественной среды обитания археолемуров; возможно также, что их истребляли как вредителей сельского хозяйства. Ареал этих приматов постепенно сокращался, пока они не исчезли окончательно — судя по всему, еще до прибытия европейцев в 1500 г.
Ссылки
Археолемуры (Википедия) перейти
Archaeolemur (Paleobiology Database) перейти
Литература
Каландадзе Н. Н., Шаповалов А. В. Судьба мегафауны наземных экосистем Мадагаскарской зоогеографической области в позднем антропогене.
Мадагаскар / Науч. ред. Б. Д. Васильев. — М.: Прогресс, 1990
Cenozoic mammals of Africa / Ed. by Werdelin L., Sanders W. J. — University of California Press, 2010
Ciochon R. L, Fleagle J. G. Primate evolution and human origins. — Hawthorne, NY: Aldine de Gruyter, 1987
Encyclopedia of Human Evolution and Prehistory / Ed. by Delson Е., Tattersall I., Van Couvering J., Brooks A. S. — Taylor & Francis, 2000
Jungers W. L., Lemelin P., Godfrey L. R., Wunderlich R. E., Burney D. A., Simons E. L., Chatrath P. S., James H. F., Randria G. F. N. The hands and feet of Archaeolemur: metrical affinities and their functional significance/ Journal of Human Evolution 49, 2005
Lemurs: Ecology and Adaptation / Ed. by Gould L., Sauther M. L. — Springer Science & Business Media, 2006
Mittermeier R. A., Louis E. E., Richardson M., Schwitzer C., Langrand O., Rylands A. B., Hawkins F., Rajaobelina S., Ratsimbazafy J., Rasoloarison R., Roos C., Kappeler P. M., MacKinnon J. Lemurs of Madagascar (3rd ed.). — Conservation International, 2010
Orlando L., Calvignac S., Schnebelen C., Douady C. J., Godfrey L. R., Hänni C. DNA from extinct giant lemurs links archaeolemurids to extant indriids. BMC Evolutionary Biology 121 (8), 2008
Ravosa M. J., Simons E. L. Mandibular growth and function in Archaeolemur. The American Journal of Physical Anthropology 95 (1), 1994
Szalay F., Delson E. Evolutionary history of the Primates. — New York: Academic Press, 1979
Tattersall I. The Primates of Madagascar. — New York: Columbia University Press, 1982
The Natural History of Madagascar / Ed. by Goodman S. M., Benstead J. P. — University of Chicago Press, 2003
The Primate Fossil Record / Ed. by Hartwig W. C. — Cambridge: Cambridge University Press, 2002