Бабакотии — Babakotia
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Суперотряд Эвархонтоглиресы (Euarchontoglires)
Грандотряд Эвархонты (Euarchonta)
Миротряд Приматообразные (Primatomorpha)
Отряд Приматы (Primates)
Подотряд Мокроносые приматы (Strepsirrhini)
Инфраотряд Лемурообразные (Lemuriformes)
Надсемейство Лемуроиды (Lemurоidеа)
Семейство †Палеопропитековые (Palaeopropithecidae)
Род †Бабакотии (Babakotia)
Вид †Babakotia radofilai
Бабакотии (Babakotia Godfrey, Simons, Chatrath et Rakotosamimanana, 1990) — монотипический род семейства палеопропитековых (Palaeopropithecidae Tattersall, 1973), происходящий из голоцена Мадагаскара.

Реконструкция внешнего облика бабакотии (Babakotia radofilai). Автор — Smokeybjb.
Единственный установленный вид — Babakotia radofilai Godfrey, Simons, Chatrath et Rakotosamimanana, 1990 (тип). Его остатки обнаружены в северной и северо-западной частях Мадагаскара (местонахождения Антсируандуха и Анджохибе). Они датируются голоценом.
История изучения
Впервые остатки бабакотии обнаружили в ходе экспедиций, организуемых с начала 1980-х гг. антропологом Э. Симонсом. Их открыли в пещере Антсируандуха горного массива Анкарана на севере Мадагаскара. Найденные остатки принадлежат по меньшей мере 4 особям, от одной из которых сохранился почти полный скелет, принадлежавший, по всей видимости, неполовозрелому животному. Типовой образец представлен фрагментом левой половины верхней челюсти с клыком и 2 резцами, правой стороной нижней челюсти с полностью сохранившимся набором предкоренных и коренных, а также диафизами плечевой и бедренной костей. Из других элементов посткраниального скелета известны локтевая кость и фрагмент лучевой, некоторые кости пясти и запястья, часть поясничных позвонков, бедренная кость, пяточная кость и др. Помимо этого, остатки бабакотии были обнаружены в пещере Анджохибе северо-западе острова, в 400 км к югу от плато Анкарана.
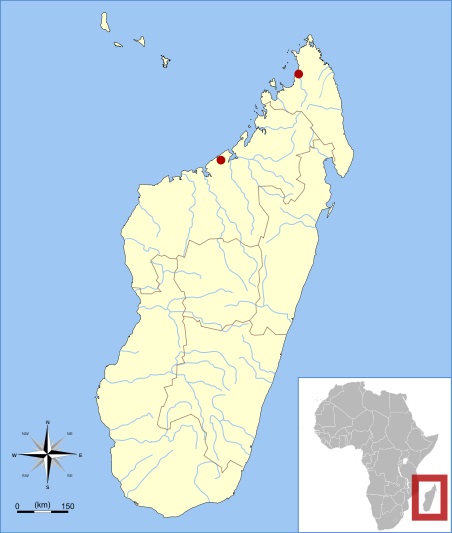
Карта местонахождений остатков бабакотии (Babakotia radofilai).
Таксон был выделен коллективом исследователей на основании особенностей строения зубной системы в 1990 г. Родовое наименование Babakotia происходит от малагасийского слова babakoto — распространенного местного названия лемура индри. Видовой эпитет radofilai был присвоен в честь выходца из Франции, математика Ж. Радофилао (Дюфло). Он проявлял живой интерес к исследованию пещер, где позже были обнаружены первые остатки бабакотии, и составил их карту.
Найденные остатки были сразу распознаны как принадлежащие близкому родственнику палеопропитеков (Palaeopropithecus) и вместе с остатками мезопропитеков (Mesopropithecus) помогли определить место палеопропитековых лемуров (Palaeopropithecidae) относительно как современных индриевых (Indriidae), так и относительно еще одного вымершего семейства — археолемуровых (Archaeolemuridae). Черепа последних по внешне сходны с черепами индриевых, но их отличаются высокоспециализированными зубами, непохожими на зубы индриевых. Напротив, бабакотию и мезопропитеков сближают с индриевыми не только очертания черепа, но и строение зубов, что дало основание утверждать о близкой родственной связи индриевых и палеопропитековых и более дальнем родстве тех и других с археолемуровыми. Это мнение остается преобладающим и сейчас, хотя существует точка зрения, что археолемуровые ближе к палеопропитековым, чем индриевые.
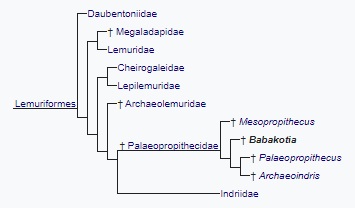
Место бабакотии (Babakotia radofilai) в систематике лемурообразных.
Кроме того, остатки бабакотии помогли установить, что палеопропитековые не передвигались, подобно современным индриевым, быстрыми прыжками с ветки на ветку, а медленно переползали по стволам и кормились, как современные ленивцы, вися горизонтально на ветвях, хотя, вероятно, и могли совершать небольшие прыжки.
Строение и функции
Бабакотия имеет средние для своего семейства размеры: длина ее черепа составляет 11,4 см, расчетная общая длина — около 75 см, вес оценивается в 15-25 кг.
Череп по своим очертаниям напоминает таковой короткохвостого индри (Indri Indri), но гораздо массивнее. Глазницы большие, направленные вперед. Надглазничные валики или другие окологлазничные утолщения отсутствуют, но имеется мощный заглазничный отросток. Скуловые дуги мощные. Саггитальный и затылочный гребни хорошо выражены. Костное небо вытянутое и узкое, практически прямоугольное, в средней части сглаженное — эта же черта, но еще более ярко выраженная, характерна также для палеопропитеков и археоиндри. Слуховой барабан увеличен, барабанное кольцо расположено внутри нее. Нижнечелюстной симфиз, как у индриевых и мезопропитеков, не срастался.
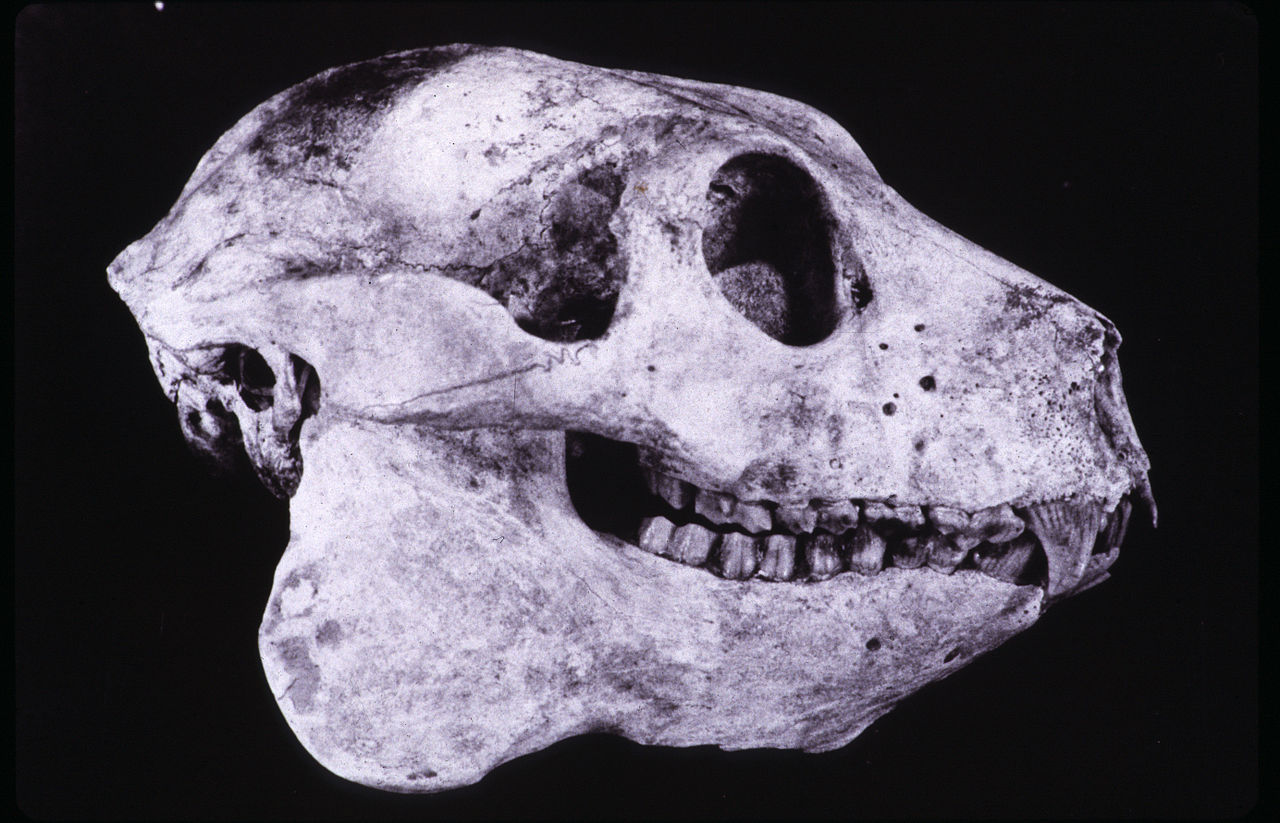
Череп с нижней челюстью бабакотии (Babakotia radofilai).
Зубная формула сходна с таковой у остальных палеопропитековых и индриевых — I2/i1, C1/c1, P2/р2, M3/m3, или I2/i2, C1/c0, P2/р2, M3/m3 (разночтения связаны с тем, что неясно до конца, является ли один из постоянных зубов резцом или клыком). Центральные верхние резцы длиннее боковых и разделены щелью. Подобно мезопропитекам, но не археоиндри и палеопропитекам, 4 нижних резца образуют гребенку, в отличие от более распространенного среди лемуров гребня из 6 резцов. По сравнению с индриевыми, верхние предкоренные удлинены. Коронка Р1 характеризуется выпуклостью в средней части, выдающейся за шейку зуба, а коронка Р2 — билофодонтией, т. е. наличием на жевательной поверхности 2 поперечных гребней, соединяющих 4 бугорка. Для коренных характерны широкая режущая кромка и мелкие зазубрины на эмали; их режущий коэффициент (показатель соотношения длины режущих кромок к общей длине коренных зубов), вообще высокий у субфоссильных лемуров, у бабакотии достигает наибольшего значения — до 50 %. Эти черты уникальны для бабакотии, отличая ее как от индриевых, так и от других палеопропитековых. Как коренные, так и предкоренные сплющены с боков, что в целом типично для индриевых и палеопропитековых.

Неполный череп бабакотии (Babakotia radofilai).
Уровень специализации бабакотии к древесному образу жизни выше, чем у мезопропитеков, но ниже, чем у палеопропитеков. Об этом свидетельствует целый ряд адаптационных изменений позвоночника, таза и конечностей. Передние конечности относительно длиннее задних и развиты лучше них. Индексы пропорций конечностей: интермембральный — 118,5 (у мезопропитеков — от 97 до 113); гумерофеморальный — 115; брахиальный — 100; круральный — 93. Лобковая кость удлинена. Нижняя ость подвздошной кости редуцирована. Седалищная ость зачаточная. Суставы бедра и лодыжки высокоподвижны. Головка бедренной кости шаровидная, лишенная ямки. Передняя большеберцовая лодыжка и пяточная кость редуцированы. Предплюсна укорочена. Большие пальцы как на кисти, так и на стопе несколько укорочены по сравнению с остальными. Проксимальные фаланги пальцев удлинены и искривлены. Имелся рудиментарный хвост.
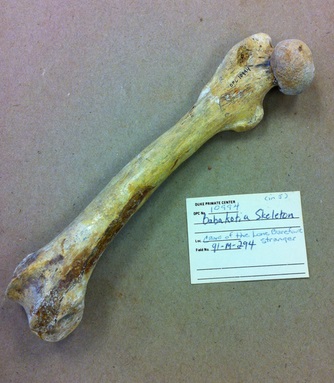
Бедренная кость бабакотии (Babakotia radofilai).
Образ жизни
Экологически бабакотия, как и другие палеопропитековые, больше всего напоминала современных ленивцев. Основную часть времени она проводила в кронах деревьев, перелезая по толстым ветвям с помощью сильных конечностей и, вероятно, подолгу висела в положении спиной вниз.
Соотношение длины плечевой и бедренной костей, составляющее почти 1,2, резко отличается от соотношения у видов, передвигающихся прыжками — у последних бедренная кость, наоборот, значительно длиннее плечевой; даже у лемуров вари (Varecia) этот показатель обратен соотношению, установленному для бабакотии. Короткие задние конечности, характерные для всех палеопропитековых, хорошо приспособлены для лазания и висения, но не для прыжков, которые совершают индриевые. Из современных млекопитающих интермембральный индекс, схожий с таковым у бабакотии, зафиксирован у ведущих преимущественно висячий образ жизни двупалых ленивцев (Choloepus), а также у горилл (Gorilla).
Анализ строения полукружных каналов внутреннего уха и позвонков поясничного отдела позволяет сделать вывод о медлительности движений, а также, в отличие от еще более специализированных палеопропитеков, об адаптации к вертикальному лазанию, как у орангутанов (Pongo) или лориевых (Loridae). На такие же выводы наталкивает больша́я длина ступней и ладоней относительно длины всей конечности. Эта черта, характерная также для палеопропитеков и еще одного вымершего рода лемуров — мегаладаписов (Megaladapis), — из ныне живущих приматов сопоставима только с таковым у орангутанов. Кости лучезапястного сустава тоже указывают на приспособленность к вертикальному лазанию.
Проксимальные фаланги пальцев удлинены и искривлены, что способствовало надежному захвату веток. Это характерно для всех палеопропитековых, но не характерно для индриевых и археолемуровых, чьи проксимальные фаланги искривлены под вдвое меньшими углами, чем у бабакотии. При этом схожую степень искривления фаланг демонстрируют ленивцы, а также коаты (Ateles) и лемуры вари (Varecia), по последним данным, также ведущие висячий образ жизни.
На основании размеров тела, строения и особенностей износа зубов можно предполагать, что бабакотия была преимущественно листоядной; вероятно, в ее рацион также входили плоды и твердые семена. Глазницы, сопоставимые по абсолютным размерам с глазницами современных дневных лемуров, а также узкий зрительный канал свидетельствуют о дневной активности и низкой резкости зрения.
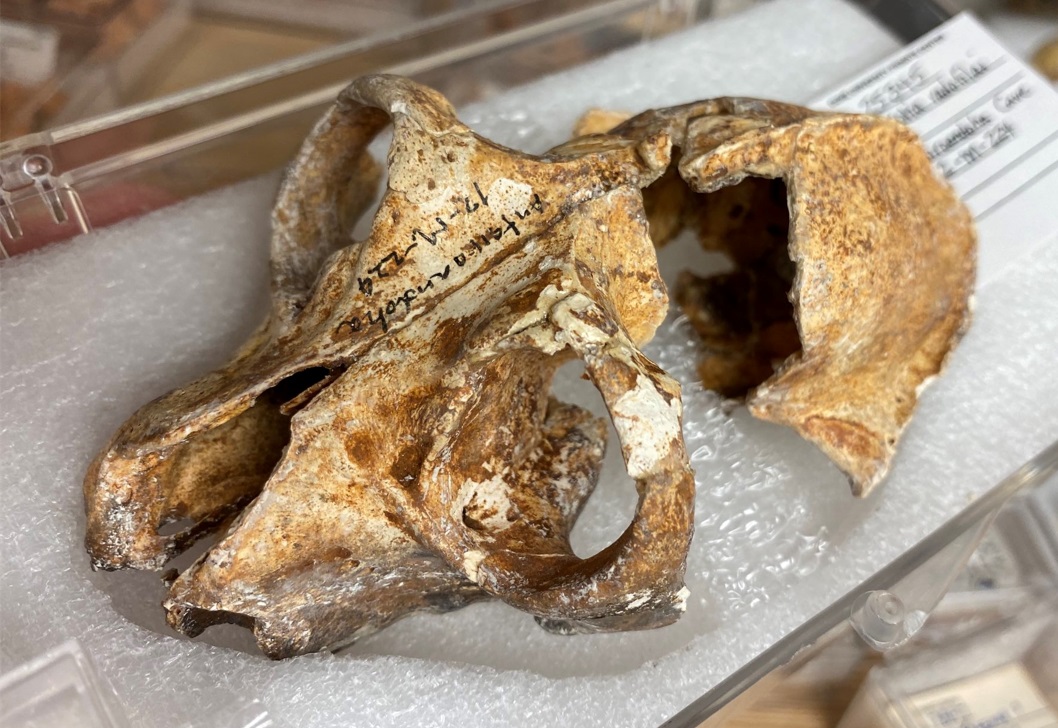
Неполный череп бабакотии (Babakotia radofilai).
Выраженная сезонность климата в районе обитания бабакотии подразумевает образ жизни и, в конечном итоге, физиологию, сопряженные с экономным расходованием энергии и максимально эффективным использованием пищевых ресурсов. В частности, для бабакотии, как и для других палеопропитековых и индриевых, характерно раннее прорезание постоянных зубов, что обеспечивало выживаемость молодых особей в первый сухой сезон после отлучения от груди. Исходя из особенностей развития зубов, предполагается, что как отнятие от груди, так и в дальнейшем достижение половой зрелости происходили, по-видимому, быстрее, чем у обезьян сопоставимых размеров.
Относительно малый объем мозга бабакотии рассматривается в качестве доказательства ее невысокой социальной активности и проживания небольшими группами (хотя специфические детали микроизноса зубной гребенки указывают на то, что она использовалась для груминга — расчесывания и чистки меха, — неизвестно, носило ли это занятие социальный характер).
Небольшая область распространения даже в период, когда почти весь Мадагаскар был покрыт лесом, может быть связана со специфическим рационом, конкурентным исключением или какими-либо другими неизвестными факторами. Тем не менее, показано, что ареал бабакотии пересекался с ареалами 2 других видов ленивцевых лемуров — Palaeopropithecus maximus и Mesopropithecus dolichobrachion. Небольшие различия в характере износа зубов у родов Babakotia и Palaeopropithecus (большее количество ямок в эмали у бабакотии) объясняются вероятным содержанием более жестких листьев либо семян в диете.
Хотя нет однозначных указаний на то, кто являлся естественнымыми врагами бабакотии, ее остатки находят в тех же местах и в тех же слоях, что и остатки крупного крокодила Voay robustus (в первую очередь на плато Анкарана). Найдены многочисленные остатки других крупных субфоссильных лемуров (Palaeopropithecus, Archaeolemur, Megaladapis) со следами крокодильих зубов. Опасность для бабакотии представляли также гигантская фосса (Cryptoprocta spelea) и крупные хищные птицы, но они в основном предпочитали более мелкую добычу.
Вымирание
Судя по всему, вымирание бабакотии связано с деятельностью человека — выжигание тропических лесов под пашни и пастбища приводило к уменьшению площади местообитания этого и многих других лемуров, а охота способствовала сокращению их популяции. Полагают, что это животное окончательно вымерло порядка 2000-1000 лет назад, т. е. незадолго до прибытия на Мадагаскар людей или вскоре после этого. Однако к настоящему времени единственная произведенная радиоуглеродная датировка его остатков составила 4400 ± 60 лет.
Ссылки
Бабакотии (Википедия) перейти
Babakotia (Paleobiology Database) перейти
Литература
Каландадзе Н. Н., Шаповалов А. В. Судьба мегафауны наземных экосистем Мадагаскарской зоогеографической области в позднем антропогене.
Ankel-Simons F. Primate Anatomy (3rd ed.). — Academic Press, 2007
Burney D. A., James H. F., Grady F. V., Rafamantanantsoa J., Ramilisonina, Wright H. T., Cowart J. B. Environmental Change, Extinction and Human Activity: Evidence from Caves in NW Madagascar. Journal of Biogeography 24, 1997
Cenozoic mammals of Africa / Ed. by Werdelin L., Sanders W. J. — University of California Press, 2010
Crowley B. E. A refined chronology of prehistoric Madagascar and the demise of the megafauna. Quaternary Science Reviews 29, 2010
Geissmann T. Vergleichende Primatologie. — Springer, 2003
Godfrey L. R., Jungers W. L. The Extinct Sloth Lemurs of Madagascar. Evolutionary Anthropology 12, 2003
Godfrey L. R., Semprebon G. M., Jungers W. L., Sutherland M. R., Simons E. L., Solounias N. Dental use wear in extinct lemurs: evidence of diet and niche differentiation. Journal of Human Evolution 47 (3), 2004
Godfrey L. R., Simons E. L., Chatrath P. J., Rakotosamimanana B. A new fossil lemur (Babakotia, Primates) from northern Madagascar. Comptes Rendus de l'Académie des Sciences 81 (2), 1990
Goodman M. S., Jungers W. L. Extinct Madagascar: Picturing the Island's Past. — The University of Chicago Press, 2014
Granatosky M. C., Miller C. E., Boyer D. M., Schmitt D. Lumbar vertebral morphology of flying, gliding, and suspensory mammals: Implications for the locomotor behavior of the subfossil lemurs Palaeopropithecus and Babakotia. Journal of Human Evolution 75, 2014
Hamrick M. W., Simons E. L., Jungers W. L. New wrist bones of the Malagasy giant subfossil lemurs. Journal of Human Evolution 38 (5), 2000
Herrera J. P., Dávalos L. M. Phylogeny and Divergence Times of Lemurs Inferred with Recent and Ancient Fossils. Systematic Biology 65 (5), 2016
Elwyn Simons: A search for origins / Ed. by Fleagle J. G., Gilbert C. C. — New York: Springer, 2008
Encyclopedia of Human Evolution and Prehistory / Ed. by Delson Е., Tattersall I., Van Couvering J., Brooks A. S. — Taylor & Francis, 2000
Jungers W. L., Godfrey L. R., Simons E. L., Chatrath P. S., Rakotosamimanana B. Phylogenetic and functional affinities of Babakotia (Primates), a fossil lemur from northern Madagascar. Proceedings of the National Academy of Sciences 88 (20), 1991
Jungers W. L., Godfrey L. R., Simons E. L., Chatrath P. S. Phalangeal curvature and positional behavior in extinct sloth lemurs (Primates, Palaeopropithecidae). Proceedings of the National Academy of Sciences 94 (22), 1997
Karanth K. P., Delefosse T., Rakotosamimanana B., Parsons T. J., Yoder A. D. Ancient DNA from giant extinct lemurs confirms single origin of Malagasy primates. Proceedings of the National Academy of Sciences of the United States of America 102 (14), 2005
Kistler L., Ratan A., Godfrey L. R., Crowley B. E., Hughes C. E., Lei R., Cui Y., Wood M. L., Muldoon K. M., Andriamialison H., McGraw J. J., Tomsho L. P., Schuster S. C., Miller W., Louis E. E., Yoder A. D., Malhi R. S., Perry G. H. Comparative and population mitogenomic analyses of Madagascar's extinct, giant 'subfossil' lemurs. Journal of Human Evolution 79 (50), 2015
Lemurs: Ecology and Adaptation / Ed. by Gould L., Sauther M. L. — Springer Science & Business Media, 2006
Marchi D., Ruff C. B., Capobianco A., Rafferty K. L., Habib M. B., Patel B. A. The locomotion of Babakotia radofilai inferred from epiphyseal and diaphyseal morphology of the humerus and femur. Journal of morphology 277 (9), 2016
Meador L. Who Ate the Subfossil Lemurs? A Taphonomic and Community Study of Raptor, Crocodylian and Carnivoran Predation of the Extinct Quaternary Lemurs of Madagascar. — University of Massachusetts Amherst, 2018
Mittermeier R. A., Louis E. E., Richardson M., Schwitzer C., Langrand O., Rylands A. B., Hawkins F., Rajaobelina S., Ratsimbazafy J., Rasoloarison R., Roos C., Kappeler P. M., MacKinnon J. Lemurs of Madagascar (3rd ed.). — Conservation International, 2010
Natural Change and Human Impact in Madagascar / Ed. by Goodman S. M., Patterson B. D. — Smithsonian Institution Press, 1997
Nowak R. M. Walker's Primates of the World. — Baltimore, MD: The Johns Hopkins University Press, 1999
Orlando L., Calvignac S., Schnebelen C., Douady C. J., Godfrey L. R., Hänni C. DNA from extinct giant lemurs links archaeolemurids to extant indriids. BMC Evolutionary Biology 121 (8), 2008
Rafferty K. L., Teaford M. F., Jungers W. L. Molar microwear of subfossil lemurs: improving the resolution of dietary inferences. Journal of Human Evolution 43 (5), 2002
Reconstructing Behavior in the Primate Fossil Record / Ed. by Plavcan J. M., Kay R. F., Jungers W. L., van Schaik C. P. — Springer Science & Business Media, 2012
Shapiro L. J., Seiffert C. V. M., Godfrey L. R., Jungers W. L., Simons E. L., Randria G. F. N. Morphometric Analysis of Lumbar Vertebrae in Extinct Malagasy Strepsirrhines. American Journal of Physical Anthropology 128 (4), 2005
Simons E. L., Burney D. A., Chatrath P. S., Godfrey L. R., Jungers W. L., Rakotosamimanana B. AMS 14C Dates for Extinct Lemurs from Caves in the Ankarana Massif, Northern Madagascar. Quaternary Research 43 (2), 1995
Simons E., Godfrey L. R., Jungers W. L., Chatrath P., Rakotosamimanana B. A New Giant Subfossil Lemur, Babakotia, and the Evolution of the Sloth Lemurs. Folia Primatologica 58 (4), 1992
Simons E. L., Godfrey L. R., Vuillaume-Randriamanantena M., Chatrath P. S., Gagnon M. Discovery of New Giant Subfossil Lemurs in the Ankarana Mountains of Northern Madagascar. Journal of Human Evolution 19 (3), 1990
The Natural History of Madagascar / Ed. by Goodman S. M., Benstead J. P. — University of Chicago Press, 2003
The Primate Fossil Record / Ed. by Hartwig W. C. — Cambridge: Cambridge University Press, 2002
Walker A., Ryan T. M., Silcox M. T., Simons E. L., Spoor F. The Semicircular Canal System and Locomotion: The Case of Extinct Lemuroids and Lorisoids. Evolutionary Anthropology 17, 2008