Биармозухи — Biarmosuchus
Класс †Синапсиды (Synapsida)
Подкласс †Терапсиды (Therapsida)
Надотряд †Динообразные (Dinomorpha)
Отряд †Диноцефалы (Dinocephalia)
Подотряд †Титанозухиды (Titanosuchida)
Надсемейство †Эотитанозухоиды (Eotitanosuchoidea)
Семейство †Эотитанозуховые (Eotitanosuchidae)
Род †Биармозухи (Biarmosuchus)
Вид †Biarmosuchus tener
Вид †Biarmosuchus tchudinovi
Биармозухи (Biarmosuchus Tchudinov, 1960) — род семейства эотитанозуховых (Eotitanosuchidae Tchudinov, 1960), происходящий из средней перми Европы. Установлены 2 вида.

Реконструкция внешнего облика молодой особи биармозуха (Biarmosuchus tener). Автор — Д. Богданов.
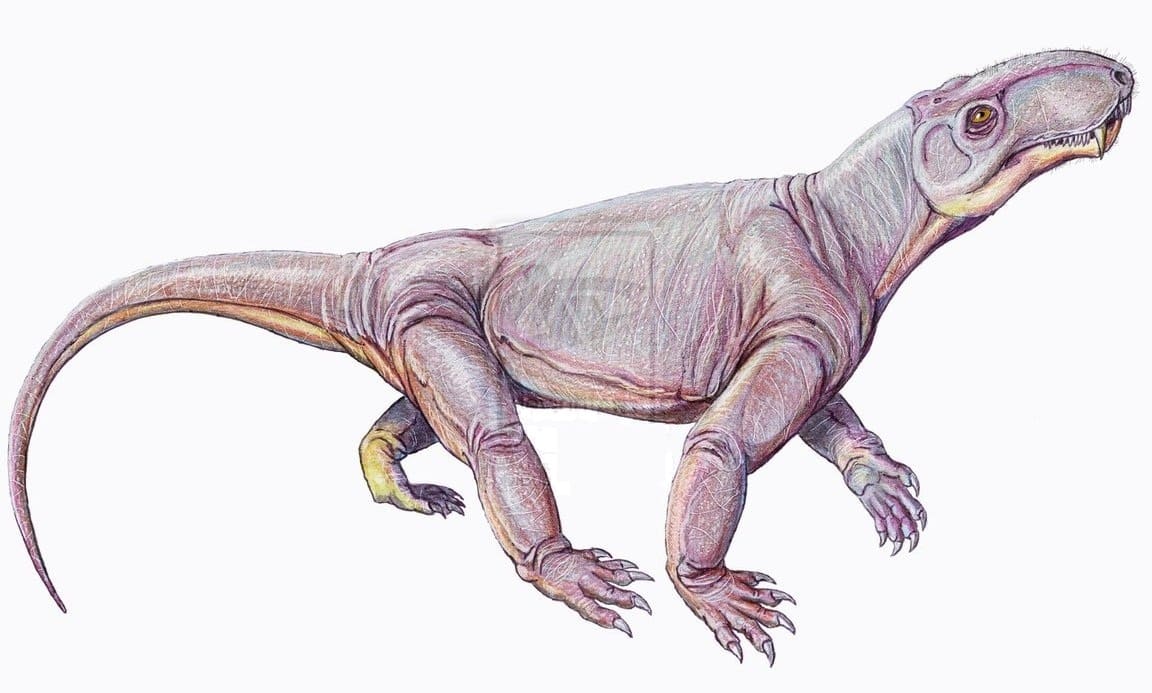
Реконструкция внешнего облика взрослой особи биармозуха (Biarmosuchus tener), или "эотитанозуха" (Eotitanosuchus olsoni). Автор — Д. Богданов.
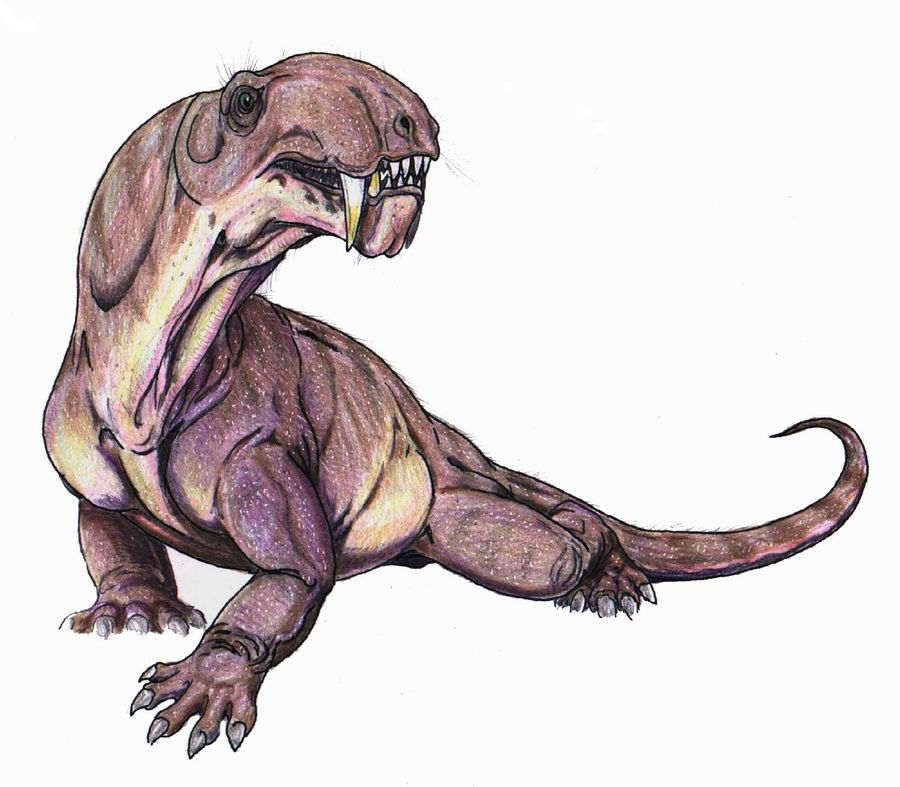
Реконструкция внешнего облика взрослой особи биармозуха (Biarmosuchus tener), или "ивантозавра" (Ivantosaurus ensifer). Автор — Д. Богданов.
Синонимы: Eotitanosuchus Tchudinov, 1960; Biarmosaurus Tchudinov, 1964; Ivantosaurus Tchudinov, 1983.
Название Biarmosuchus происходит от Биармии — исторической области на севере Восточной Европы, предположительно располагавшейся на территории современной Архангельской области, в бассейне Северной Двины. Название рода Eotitanosuchus переводится как «ранний крокодил-титан». Ivantosaurus, или "Ивана Антоновича ящер", назван в честь российского палеонтолога И.А. Ефремова.
Biarmosuchus tener Tchudinov, 1960 (тип) — типовой вид, известный из местонахождения Ежово (Очер) (окрестности деревни Ежово, Очерский район, Пермская область, Россия). Они датируются средней пермью (уржумский горизонт, уржумский ярус, биармийский отдел). Синонимы: Eotitanosuchus olsoni Tchudinov, 1960; Biarmosaurus antecessor Tchudinov, 1964; Ivantosaurus ensifer Tchudinov, 1983; Ivantosaurus signifer Thudinov in litt, err.
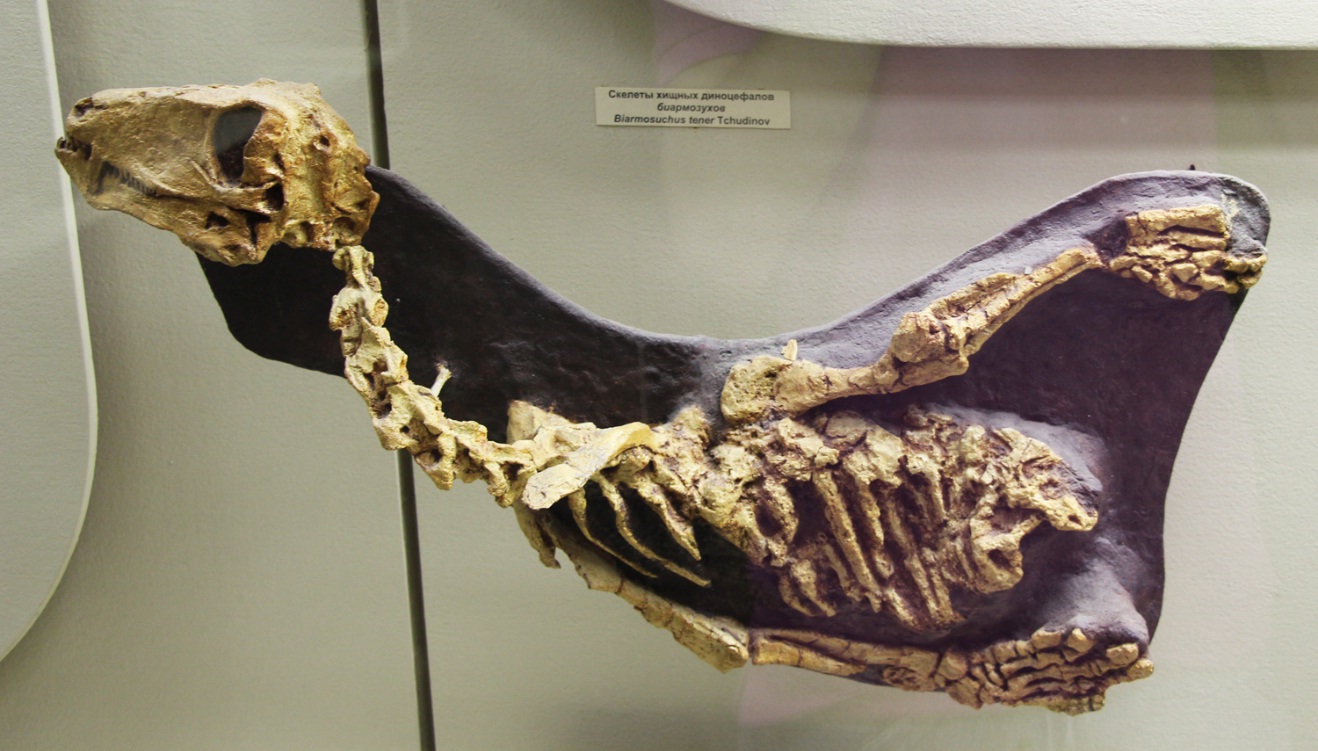
Скелет в породе молодой особи биармозуха (Biarmosuchus tener) из местонахождения Ежово, Пермская область, Россия.
Таксоны Biarmosuchus tener и Eotitanosuchus olsoni были описаны П.К. Чудиновым в 1960 г. в одной и той же статье в составе семейства Eotitanosuchidae, установленного там же. Название Eotitanosuchus olsoni имеет только то преимущество, что соответствует наименованию семейства, однако типовой экземпляр единственного вида крайне неполон (череп без части неба и нижней челюсти). В то же время, типовой экземпляр Biarmosuchus tener — почти полный (без передних конечностей) скелет с полным черепом, и потому было решено считать старшим синонимом название Biarmosuchus tener.



Реконструированный скелет молодой особи биармозуха (Biarmosuchus tener) из местонахождения Ежово, Пермская область, Россия.
В 1964 г. П.К. Чудиновым из одновозрастных отложений Ежово по черепу и костям таза был описан Biarmosaurus antecessor. Позднее сам автор синонимизировал его с Biarmosuchus tener. В 1983 г. П.К. Чудиновым все из того же слоя Ежовского местонахождения по обеим максиллам и квадратной кости был описан Ivantosaurus ensifer. Проведенный М.Ф. Ивахненко сравнительный анализ строения черепа Biarmosuchus tener, Eotitanosuchus olsoni и Ivantosaurus ensifer не выявил существенных различий между ними, за исключением гораздо более крупных размеров двух последних форм. Имеющиеся небольшие морфологические отличия, по мнению М.Ф. Ивахненко, являются следствием онтогенетических изменений, т. е. Eotitanosuchus olsoni и Ivantosaurus ensifer являются взрослыми особями Biarmosuchus tener. Синонимичность Eotitanosuchus olsoni и Ivantosaurus ensifer сегодня практически не вызывает сомнений, но отнесение их к одному роду с Biarmosuchus tener оспаривается многими исследователями.
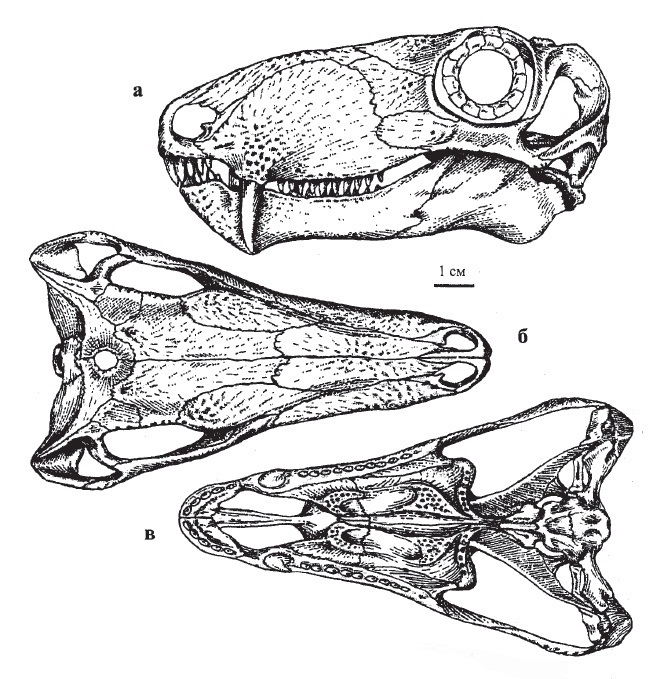
Прорисовки черепа молодой особи биармозуха (Biarmosuchus tener) из местонахождения Ежово, Пермская область, Россия: сбоку (а), сверху (б) и снизу (в).
Молодые экземпляры типового вида биармозухов достигали около 1,5 м в длину, общая длина взрослых особей оценивается в 3-4 м.
Череп довольно высокий, клиновидный, со слабо суженным лицевым отделом. Предглазничная часть черепа удлиненная, заглазничная — короткая. Морда оканчивается круто. Ноздри большие, сближенные друг с другом. Предклыковый край верхней челюсти сильно скошен вверх. Зубной край максиллы выпуклый и сильно отогнутый вниз по отношению к уровню предчелюстной и скуловой костей. Восходящие пластины премаксилл очень сильно вдаются между носовыми костями. Глазница большая (особенно у молодых особей), продольно-овальная, с развитым кольцом склеротики. Верхний край заглазничной кости образует утолщенный нависающий выступ. Височное окно слабо открытое сверху, достигающее по горизонтальному диаметру 0,75 диаметра глазницы при максимальной высоте черепа в скуловой области не менее 7 см. У взрослых особей височное окно относительно более расширено в верхней части.
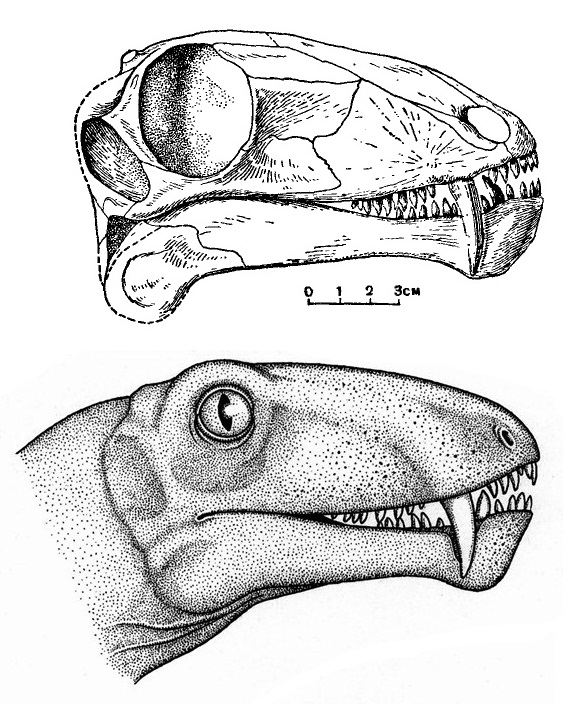
Прорисовка черепа молодой особи биармозуха (Biarmosuchus tener), или "биармозавра" (Biarmosaurus antecessor), из местонахождения Ежово, Пермская область, Россия, и его портретная реконструкция (автор — А. Кац).
Крыша межглазничной и теменной областей тонкая. Теменная область широкая, сильно вогнутая. Теменные кости очень короткие, особенно у взрослых особей, так что пенальное отверстие относительно их выглядит очень крупным. Пинеальное отверстие окружено невысоким костным валиком и расположено на теменном бугре. Канал пинеального органа продольноовальный, направленный назад и вверх. Теменной бугор сравнительно невысокий и расположен у края затылка; задняя часть его переходит на затылочную поверхность черепа и образована теменными костями. Пахиостозные утолщение и бугристые разрастания костей крыши черепа отсутствуют. Затылок слабо наклонен верхней частью вперед. Заднетеменная кость широкая, спускающаяся выпуклым полуконусом вниз и оканчивающаяся у большого затылочного отверстия. Квадратные кости длинные, ориентированные вертикально. По крайней мере у молодых особей наружный мыщелок квадратной кости расположен ниже внутреннего. Таблитчатые и верхняя затылочная кости очень высокие и сходные по форме. Околозатылочные отростки длинные, массивные и сильно отогнуты вниз.
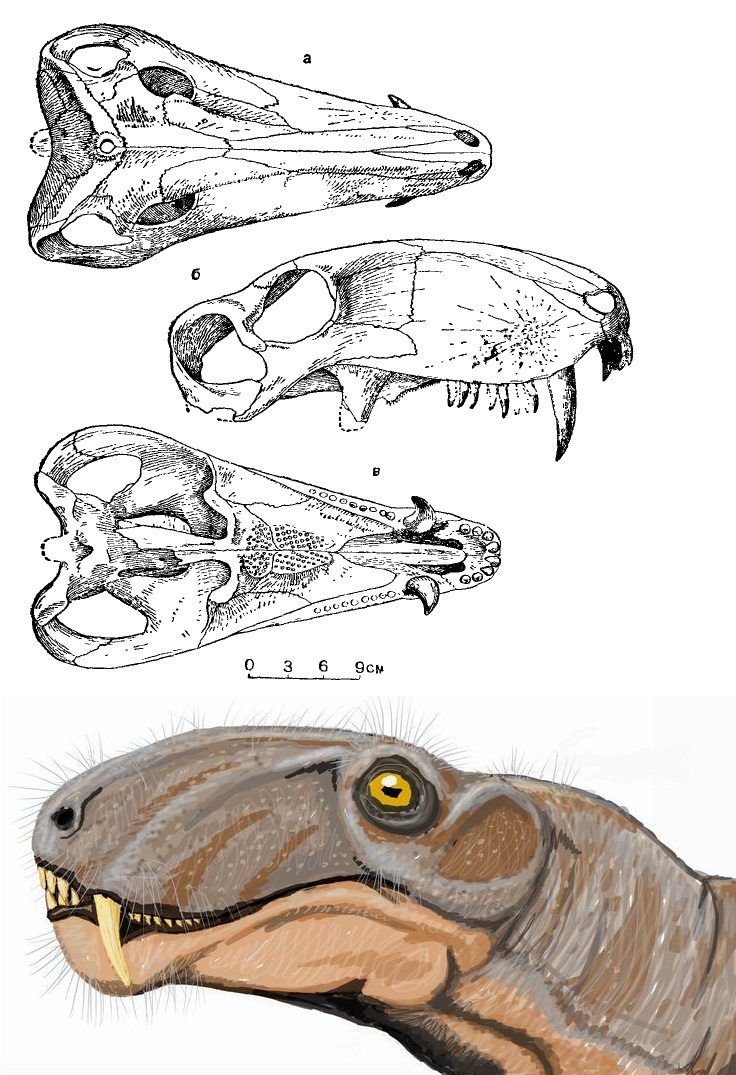
Череп взрослой особи биармозуха (Biarmosuchus tener), или "эотитанозуха" (Eotitanosuchus olsoni), из местонахождения Ежово, Пермская область, Россия, и его портретная реконструкция (автор — Д. Богданов).
Передние участки хоан расширены полукруглыми вырезками в премаксиллах. Задние края хоан находятся на уровне II-х заклыковых зубов. Поперечные ветви крыловидных костей очень массивные, высокие и сближенные. Озубленные небно-птеригоидные бугры не развиты. Некрупные зубы широко покрывают передние и, по-видимому, поперечные отростки птеригоидов, причем зубы на птеригоидных флангах никак не выделяются по размерам. Челюстной сустав несколько сдвинут вперед. Стремя представляет собой крупную поперечно ориентированную кость с очень большим стапедиальным отверстием. Нижняя челюсть высокая, тонкая, с хорошо выраженным подбородочным выступом и высоким, но слабым симфизом. Отогнутая пластинка угловой кости отчетливо отделена от остальной части этой кости и несет радиальные гребни.
Верхний клык увеличенный, длинный и довольно сильно сжатый с боков, с округлым передним и режущими зазубренным задним краями. Конец клыка достигает нижнего края подбородочного выступа, но не выступает за его пределы. От резцов и заклыковых зубов клык отделен отчетливыми диастемами. Соотношение длины клыка и его максимального диаметра, не изменяющееся в онтогенезе, составляет порядка 3 : 1. При возрастном увеличении размеров клыка узкая и высокая клыковая сумка приобретает двухвершинный вид, так как образуется расширение для одного сменного клыка. (У других саблезубых хищников, горгонопид, еще большее увеличение клыков достигалось другим путем — расширением клыковой сумки. При этом в сумке находился не 1, а 2-3 сменных клыка.)

Модель черепа взрослой особи биармозуха (Biarmosuchus tener), или "ивантозавра" (Ivantosaurus ensifer), из местонахождения Ежово, Пермская область, Россия, частично реконструированного на основе черепа "эотитанозуха" (Eotitanosuchus olsoni). Длина верхнего клыка преувеличена (его верхушка неизвестна) — сейчас полагают, что он выступал из максиллы примерно на 15 см.
Остальные зубы мелкие. Резцы загнутые, округлые в сечении, со слабо вздутыми наружными стенками коронок и режущими кантами. Заклыковые зубы с продольными основаниями, сжатыми с боков и смещенными к заднему краю вершинками. Передние края заклыковых зубов закругленные, задние — приостренные, мелко зазубренные. Число зубов на прмаксилле колеблется от 4 до 6, заклыковых зубов в максилле — от 9 до 13. Иногда число зубов резко отличается не только у разных особей, но и на разных сторонах одного черепа. Это связано со сменой зубов, регулярно происходившей в течение жизни животного. Дело в том, что в челюстях одновременно находились рабочие зубы, ровно расположенные в альвеолах, и частично мацерированные сменные зубы, длинные корни которых не позволяли им окончательно выпасть, но они располагались косо и беспорядочно. В частично отпрепарированном черепе не выпавшие сменные зубы выглядят как дополнительные. Таким образом, в премаксилле располагались 4 рабочих зуба, а в заклыковом ряду максиллы — 9-10. В нижней челюсти резцов 5.
Голотип Biarmosuchus tener принадлежит самой молодой найденной особи (длина черепа 16,5 см), остальные выстраиваются в ряд: голотип Biarmosuchus antecessor и еще один череп — по 20 см, следующие 2 черепа — по 24 см, голотип Eotitanosuchus olsoni — 34,5 см, голотип Ivantosaurus ensifer — реконструируемая длина 53-55 см, второй, более крупный фрагментарый череп (затылочная часть и мозговая коробка) — реконструируемая длина составляет порядка 70 см. При этом заметно меняется форма черепа — от относительно короткого, низкого, с крупными глазницами и короткими клыками к удлиненному черепу, высокому в предглазничной части, с относительно меньшими глазницами и очень длинными клыками. При увеличении длины черепа в 2 раза длина клыка и высота верхнечелюстной области увеличивается более чем в 2,5 раза, в то время как диаметр глазницы увеличивается всего лишь в 1,5 раза; предглазничный район удлиняется при этом более чем в 2 раза, но за счет относительного уменьшения диаметра глазницы идет непропорциональное удлинение височной области и височного окна (примерно в 2,5 раза).

Черепной материал биармозуха (Biarmosuchus tener) различного индивидуального возраста.
В строении черепа отмечаются черты сходства одновременно со сфенакодонтными пеликозаврами и примитивными горгонопами. Височная мускулатура связана в основном с внутренней черепной поверхностью. Верхневисочная кость утрачена. Затылок слегка наклонен антеровентрально, т. е. в направлении, обратном по сравнению с пеликозаврами, но имеет сходную пластиновидную форму. Клыки сильно выдаются, но передней «ступеньки» максиллы, как у Dimetrodon, нет. Как и у продвинутых сфенакодонтид, она простирается дорсально до носовой кости, отделяя слезную кость от ноздри. В отличие от всех пеликозавров септомаксилла выглядит с поверхности черепа длинным элементом. На черепе Eotitanosuchus olsoni сохранились отпечатки кожных покровов, лишенных чешуи.
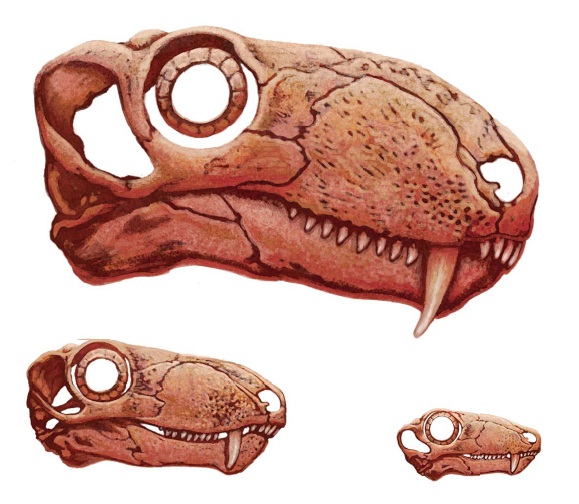
Возрастные изменения черепа биармозуха (Biarmosuchus tener). Автор — А. Атучин.
Небо сохраняет большинство примитивных признаков сфенакодонтид, включая наличие мелких зубов на небных костях и поперечных флангах крыловидных костей, но сошники заглубляются выше уровня хоан и частично сливаются. Небные и передняя часть крыловидных костей выгибаются вверх над средней линией, что наводит на мысль о существовании узкого воздушного прохода над остальным небом.
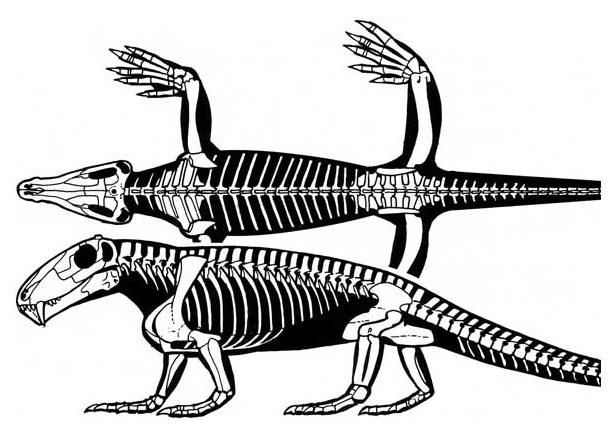
Прорисовки реконструированного скелета молодой особи биармозуха (Biarmosuchus tener), сверху и сбоку. Автор — Д. Петерс.
Посткраниальный скелет известен лишь для молодых особей, у которых он довольно грацильный. Имеется 7 шейных, 20 спинных и 3 крестцовых позвонка. Позвонки напоминают таковые примитивных сфенакодонтных пеликозавров, без увеличенных остистых отростков. Туловищный отдел позвоночника относительно короткий. Крестцовые позвонки несросшиеся.
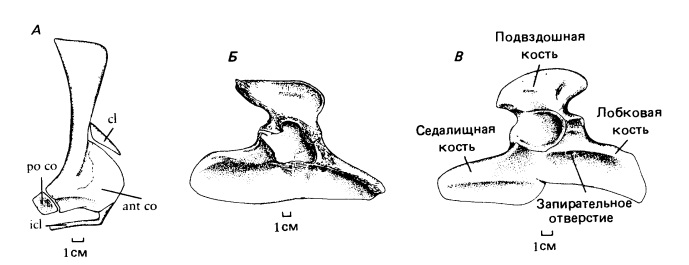
Прорисовки плечевого пояса биармозуха (Biarmosuchus tener) (А), плечевого пояса диметродона (Dimetrodon sp.) (Б) и тазового пояса биармозуха (Biarmosuchus tener) (В).
Удлиненные конечности наряду со строением их поясов говорят о гораздо более прогрессивной постановке тела по сравнению с пеликозаврами. Плечевой пояс приобретает подвижность. Пластина лопатки значительно уже, чем у пеликозавров, но ключица и межключица остаются крупными элементами. Плечевой сустав утрачивает винтообразную форму, характерную для пеликозавров. Очертания таза остаются примитивными из-за обособления пластинчатых лобковой и седалищной костей. Лонная область широко открыта спереди. Лобковые кости соединены в вогнутом симфизе. Седалищная кость короткая и высокая. Гленоидная и вертлужная впадины ориентированы более вентрально, и бедренная кость приближается к парасагиттальному положению, как у умеренно продвинутых текодонтов.
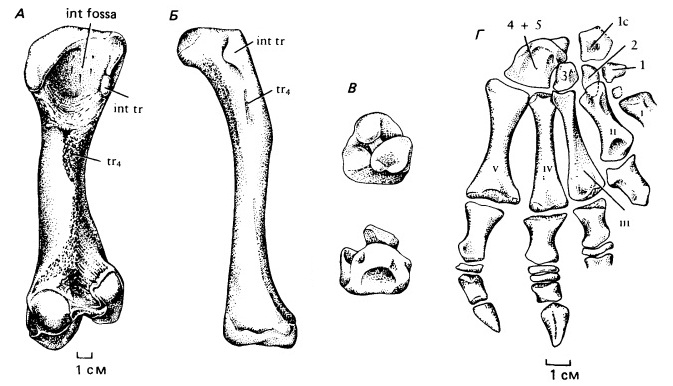
Прорисовки вентральных видов бедренной кости диметродона (Dimetrodon sp.) (А) и биармозуха (Biarmosuchus tener) (Б). В — прорисовки медиального и латерального вида сложной таранной кости биармозуха (Biarmosuchus tener). Г — прорисовка скелета стопы биармозуха (Biarmosuchus tener). Сокращения: int fossa — межвертлужная яма (fossa intertrochanterica); int tr — внутренний вертлуг (trochanter internus); lc — латеральная централия; tr4 — IV-й вертлуг; 1-5 — дистальные тарсалии; I-V — метатарсалии.
Плечевая кость сохраняет примитивную форму с сильно расширенными концами, но бедренная S-образно изогнута и ее головка повернута внутрь, как у крокодилов. Широкие кисть и стопа более симметричны, чем у пеликозавров; это свидетельствует о том, что они были направлены почти прямо вперед на протяжении всего шага. Длина некоторых фаланг сильно уменьшена, предвосхищая редукцию их числа, типичную для более поздних терапсид и млекопитающих. «Избыточные» фаланги III-го и IV-го пальцев становятся дисковидными. Пальцы снабжены длинными когтями. Подобное строение конечностей могло быть связано с обитанием на заболоченных берегах с топким грунтом.
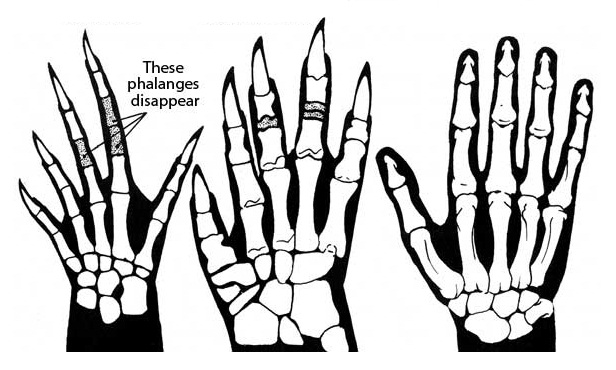
Прорисовки скелета правой кисти гаптода (Haptodus sp.), биармозуха (Biarmosuchus tener) и человека (Homo sapiens). Автор — Д. Петерс.
В целом Biarmosuchus tener по строению тела напоминал более поздних горгонопсов, но был примитивнее и крупнее (если считать Eotitanosuchus olsoni и Ivantosaurus ensifer синонимами типового вида биармозухов).
Типовый вид биармозуха является характерным представителем очерского подкомплекса очерского фаунистического комплекса наземных позвоночных. Его ископаемые остатки не редки и составляют в Ежовском местонахождении до 10 % всех находок, включая черепа разной степени сохранности и несколько более или менее полных скелетов.
Местообитаниями Biarmosuchus tener являлись заболоченные тропические леса. Его пищевая спицеализация, по-видимому, менялась по мере роста. Молодые некрупные особи, вероятно, выполняли роль очень подвижных неспециализированных прибрежных полуводных или относительно наземных охотников на мелкую добычу, а также всеядов. Крупные и тяжелые взрослые особи могли являться облигатно водными хорошо плавающими хищниками, приспособленными к питанию животными, сопоставимыми по размерам с ними самими. Увеличенные саблевидные клыки взрослого биармозуха явно служили для пробивания шкуры очень крупных растительноядных. В Очерском комплексе такой добычей могли служить эстемменозухи (Estemmenosuchus). Крупные глазницы могут свидетельствовать о сумеречном образе жизни.

Биармозух (Biarmosuchus tener) и стадо эстемменозухов (Estemmenosuchus mirabilis). Автор — А. Атучин.
Biarmosuchus tchudinovi Ivachnenko, 1999 — вид, известный из средней перми (верхнеказанский подъярус или уржумский ярус, биармийский отдел) местонахождения Сокол (Завьяловский район, Удмуртия, Россия). Синоним: Biarmosuchus sp., determ. Ivachnenko et al., 1997. Обнаружены левая максилла с частью небной костью, а также правая максилла, возможно, принадлежавшие одной особи. Расчетная длина черепа составляет порядка 20-25 см. Костная хоана длинная, продолжается назад почти до уровня середины заклыкового зубного ряда. По сравнению с типовым видом, верхнечелюстной клык более длинный и тонкий: соотношение ширины его основания и длины составляет 1 : 5.
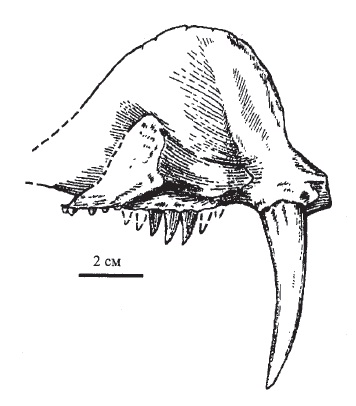
Прорисовка левой верхнечелюстной кости с частью небной биармозуха (Biarmosuchus tchudinovi) из местонахождения Сокол, Удмуртия, Россия; изнутри.
Исчезновение биармозухов, вероятно, связано с экологическим кризисом, вызванным некоторым иссушением климата. Им на смену пришли более прогрессивные хищные диноцефалы антеозавроиды.
Ссылки
Биармозух (Википедия) перейти
Ивантозавр (Википедия) перейти
Эотитанозух (Википедия) перейти
Biarmosuchus (Paleobiology Database) перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Голубев В. К. Стратотипы и опорные разрезы верхней перми Поволжья и Прикамья. — Казань: Экоцентр, 1996
Иванов А. О., Черепанов Г. О. Ископаемые высшие позвоночные. — СПб.: Издательство Санкт-Петербургского университета, 2001
Ивахненко М. Ф. Биармозухи Очерского фаунистического комплекса Восточной Европы. Палеонтологический журнал 3, 1999
Ивахненко М. Ф. Тетраподы Восточно-Европейского плаката — позднепалеозойского территориально-природного комплекса. — Пермь, 2001
Ивахненко М. Ф. Эволюция позднепалеозойских тетрапод как эволюция их биоморф. Рожнов С.В. (ред.). Эволюция биосферы и биоразнообразия. — М.: Товарищество научного издания КМК, 2006
Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1 / Отв. ред. Ивахненко М. Ф. и Курочкин Е. Н. — М.: ГЕОС, 2008
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Орлов Ю. А. В мире древних животных. Очерки по палеонтологии позвоночных. — М.: Наука, 1989
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГЕОС, 2009