Брахипотерии — Brachypotherium
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Клада Скротиферы (Scrotifera)
Клада Феревунгуляты (Fereuungulata)
Клада Эвунгуляты (Euungulata)
Клада Мезаксонии (Mesaxonia)
Отряд Непарнопалые (Perissodactyla)
Подотряд Тапирообразные (Tapiromorpha)
Инфраотряд Носорогообразные (Ceratomorpha)
Надемейство Риноцеротоиды (Rhinocerotoidea)
Семейство Носороговые (Rhinocerotidae)
Подсемейство †Ацератериины (Aceratheriinae)
Триба †Телеоцератини (Teleoceratini)
Род †Брахипотерии (Brachypotherium)
Вид †Brachypotherium fatehjangense
? Вид †Brachypotherium gajense
Вид †Brachypotherium pugnator
Вид †Brachypotherium perimense
? Вид †Brachypotherium аurelianense
Вид †Brachypotherium brachypus
? Вид †Brachypotherium stehlini
Вид †Brachypotherium goldfussi
Вид †Brachypotherium snowi
Вид †Brachypotherium heinzelini
Вид †Brachypotherium minor
Вид †Brachypotherium lewisi
Брахипотерии (Brachypotherium Roger, 1904) — род подсемейства ацератериин (Aceratheriinae Dollo, 1885), происходящий из позднего олигоцена - позднего миоцена Азии, раннего - позднего миоцена / ? раннего плиоцена Европы и раннего миоцена - раннего / ? позднего плиоцена Африки. Установлены 9-12 видов.

Реконструкция внешнего облика брахипотерия (Brachypotherium lewisi). Автор — О. Марцун.
Синонимы: Rhinoceros Linnaeus, 1758 (частью); Aceratherium Cope, 1873 (частью); Teleoceras Hatcher, 1894 (частью); Aprotodon Forster-Cooper, 1915 (частью); Chilotherium Ringström, 1924 (частью).
История изучения
Род Brachypotherium был выделен в 1904 г. Роджером на основании находок близ города Эппельсгейма в Германии, откуда ранее было описано хоботное Deinotherium. Однако еще раньше он использовал это название в неопубликованной рукописи. Оно образовано древнегреческими словами βραχύς (brachýs) — "короткий", πούς (pous) — "нога" и θηρίον (thērion) — "зверь", что указывает на укороченные конечности этих животных. Протеро (1998 г.) отнес данный род к трибе Teleoceratini.

Неполный череп и нижняя челюсть брахипотерия (Brachypotherium aurelianense).
Строение и функции
Брахипотерии представляли собой крупных носорогов: у разных их видов длина черепа составляет 45-71 см, общая длина тела — 2,6-3,5 м, высота в холке — 1,2-1,5 м, вес варьировал в пределах 1,5-3 т.



Смонтированный скелет брахипотерия (Brachypotherium aurelianense var. gailiti) из среднего миоцена среднего течения реки Джиланчик в центральной части Казахстана.
Череп сравнительно короткий и широкий, клинообразно суживающийся кпереди. Носовые кости таже укорочены, прямые, узкие и суженные к переднему концу. Носовые кости не соединяются с предчелюстными, заканчиваясь далеко от их переднего края; костная носовая перегородка отсутствует. Судя по мозолистому утолщению на передней части носовых костей и следующие за ним шероховатости, особи некоторых видов (возможно, только самцы) могли обладать одним или двумя небольшими рогами, расположенными друг за другом, остальные же были безрогими. Лобные кости гладкие. Глазницы смещены довольно далеко вперед, их передний край расположен на уровне М2. Верхний профиль черепа относительно прямой. Теменные гребни иногда сближены. Затылочный отдел относительно широкий, низкий, частично закругленный в задней части и вертикально опускающийся вниз. Заслуховой и засуставный отростки слиты. Нижняя челюсть очень массивная и высокая, с широкими суставными концами. Симфиз нижней челюсти спереди несколько расширен, угол слабо выступает назад.
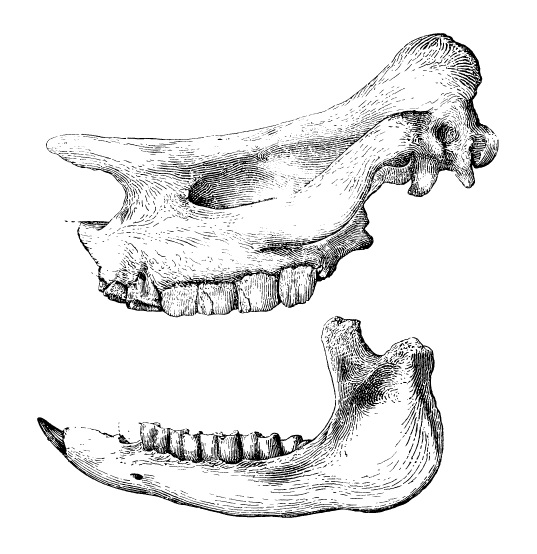
Прорисовки черепа и нижней челюсти брахипотерия (Brachypotherium aurelianense var. gailiti) из среднего миоцена среднего течения реки Джиланчик, Казахстан. Передняя часть черепа отломлена.
Зубная формула: I1/i1, C0/c0, P4/p4, M3/m3. Имеется по одной паре увеличенных верхних и нижних резцов (І1 и i2). Нижние резцы бивнеообразные. Их коронки длинные (до 11 см), конические и слегка изогнутые, так что кончики направлены вперед и вверх. Однако корни резцов относительно короткие. Клыки отсутствуют. Между резцами и щечными зубами на верхней и нижней челюсти имеются диастемы. Щечные зубы низкокоронковые, но иногда достигают средней высоты. Щечные зубы увеличиваются в заднем направлении, самыми крупными из них являются 2 последних коренных. Жевательная поверхность щечных зубов относительно широкая и устроена относительно просто, цемент на ней не развит. Предкоренные в значительной степени моляризованы. Парастильная складка на верхних коренных глубокая; кроше слабое, криста не развита.

Прорисовка жевательной поверхности правых Р1-М3 брахипотерия (Brachypotherium aurelianense var. gailiti) из среднего миоцена среднего течения реки Джиланчик, Казахстан.
Телесные пропорции бегемотоподобные. Приземистое бочкообразное туловище покоилось на укороченных тумбообразных ногах. Передние и задние конечности примерно одинаковой длины. Кисть четырехпалая, ее IV-й палец сильно укорочен. Стопа трехпалая. В конечностях наиболее развит центральный III-й метаподий. В отличие от других функционально трехпалых носорогов, II-й метаподий почти не уменьшен в размерах. В целом скелет довольно массивен.

Брахипотерий (Brachypotherium aurelianense) в естественном окружении.
Сравнение
Внешне представители рода Brachypotherium напоминали североамериканских Teleoceras, однако обладали более длинными конечностями (так, их II-й метаподий относительно длиннее). Между этими двумя родами имелись и другие отличия. Например, у Brachypotherium лобная кость имеет наибольшую ширину над передней частью глазницы; слезная железа находится впереди подглазничного отверстия базилярный бугор на клиновидной кости небольшой; забарабанный отросток тонкий и правильной формы, соприкасается с засуставным отростком; овальное отверстие отделено от рваного; М2 не намного длиннее М3. Конечности и общее сложение массивнее, чем у Diaceratherium.
Происхождение и эволюция
Род Brachypotherium состоит в подсемействе Aceratheriinae, включающем большей частью безрогих носорогов. В его рамках род принадлежит к трибе Teleoceratini, члены которой характеризуются укороченными конечностями. При этом Brachypotherium представляет довольно консервативную ветвь трибы, поскольку данный род не претерпел какой-либо значительной специализации к питанию твердой растительной пищей, несмотря на распространение открытых ландшафтов, и, таким образом, в значительной степени сохранил свои преимущественно низкокоронковые щечные зубы.
Происхождение рода Brachypotherium остается невыясненным. Древнейшие бесспорно принадлежащие ему остатки происходят из раннего миоцена Южной Азии и Европы, однако от каких более примитивных носорогов произошел Brachypotherium неизвестно. Судя по всему, его эволюционная линия рано ответвилась от линии Aceratherium. Из более молодых евроазиатских родов к Brachypotherium наиболее близки Prosantorhinus и Diаceratherium. К последнему нередко причисляется вид Brachypotherium аurelianense, который в этом случае именуется Diаceratherium аurelianense.
Ближайшим родственником и, вероятно, прямым потомком рода Brachypotherium является Teleoceras, сходный с ними морфологически и по образу жизни. Этих коротконогих носорогов часто объединяют в один род — Teleoceras или Brachypotherium, в зависимости от точки зрения авторов. Также в состав рассматриваемого рода раньше включали вид Brachypotherium аmericanum из раннего миоцена Северной Америки, который сегодня принимается в качестве Teleoceras аmericanum.
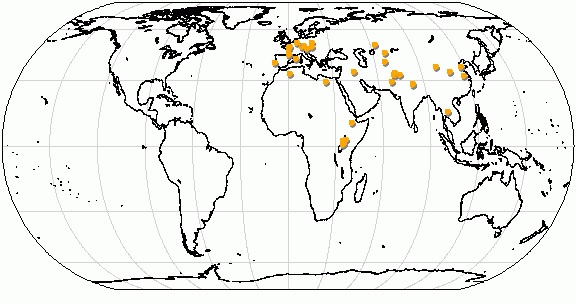
Некоторые местонахождения ископаемых остатков Brachypotherium.
Наиболее многочисленными брахипотерии были на территории Евразии — здесь их ареал простирался от Португалии и Восточного Средиземноморья до Дальнего Востока и Таиланда включительно. На севере материка эти носороги не обнаружены. Вторая группа видов получила распространение в Африке. Примечательно, что африканские брахипотерии, наряду с некоторыми ацератериями, стали первыми носорогами, заселившими эту часть света. Они иммигрировали сюда в первой половине миоцена, после чего их африканская ветвь эволюционировала параллельно евроазиатской. Некоторые местные брахипотерии превосходили по величине всех других африканских носорогов, как вымерших, так и современных. На территории Африки большинство находок сосредоточено на севере и востоке, однако имеются они и в центральной части континента. В большинстве случаев находки одиночны и фрагментарны.
Получив максимально широкое распространение в позднемиоценовое время, в раннем плиоцене брахипотерии исчезают. Одни из последних известных европейских представителей рода жили около 8 млн. лет назад на территории Болгарии, а вообще самые молодые их остатки пироисходят из Восточной Африки и датируются приблизительно в 4,2 млн. лет.
Состав
На сегодняшний день валидными считаются до 13 видов брахипотериев, некоторые из них спорны.
Brachypotherium fatehjangense (Pilgrim, 1912) — вид, известный из позднего олигоцена - позднего миоцена Пакистана (формация Чинджи и Нагри, провинции Синд и Пенджаб) и Бирмы (Чаунгтха). Описан по фрагменту верхней челюсти из Фатехянга на территории современного Пенджаба. Первоначально данный вид был описан в качестве представителя рода Chiloterium (Пилгрим, 1910 г.), затем был переопределен как Teleoceras (Пилгрим, 1912 г.), а в дальнейшем ряд исследователей относил этот вид к роду Aprotodon (Хейссиг, 1972 г.; Цю и Се, 1997 г.; Дэн, 2006 г.). Синонимы: Chiloterium fatehjangense Pilgrim, 1910; Teleoceras fatehjangense Pilgrim, 1912; Aprotodon fatehjangense Qiu and Xie, 1997.
Размеры тела относительно небольшие. Небольшая ширина щечных зубов и некоторые черты строения их жевательной поверхности указывают на питание мягким растительным кормом — например, листьями деревьев и кустарников. Особенности захоронения ископаемых остатков свидетельствуют о том, что этот носорог жил в лесах, придерживаясь относительно сухих стаций.
Brachypotherium gajense (Pilgrim, 1912) Antoine et al., 2010 — сомнительный малоизученный вид, известный по фрагментарному материалу из позднего олигоцена / раннего миоцена (хаттский ярус) Пакистана. Синоним: Aceratherium gajense Pilgrim, 1912.
Brachypotherium pugnator (Matsumoto, 1921) — вид, известный из раннего миоцена (бурдигальский ярус) Японии. Описан по изолированным зубам из отложений формации Хирамаки в регионе Тюбу на острове Хонсю. Синонимы: Teleoceras pugnator Matsumoto, 1921; Teleoceras? kaniensis Tokunaga, 1926; Teleoceras? tokiensis Tokunaga, 1926; Chilotherium pugnator (Matsumoto, 1921).

Часть нижней челюсти брахипотерия (Brachypotherium pugnator).
Brachypotherium perimense (Falconer et Cautley, 1847) Heissig, 1972 — вид, известный из раннего - позднего миоцена (бурдигальский - тортонский ярусы) Ирака, Пакистана (провинция Пенджаб), Индии (штаты Гуджарат и Пенджаб, союзная территория Джамму и Кашмир), Непала (формация Манчар, долина Синд и Данг) и Таиланда (Накорн-Ратчасим).
Крупный представитель рода. В отличие от Brachypotherium fatehjangense, имел относительно более широкие щечные зубы, что говорит о питании жесткой растительной пищей — травянистыми растениями, их корневищами и т. п. Данный вид населял в основном заболоченные местности.
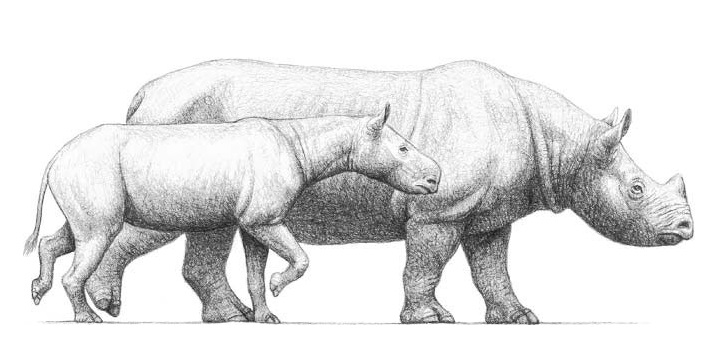
Реконструкции внешнего облика носорогов протацератерия (Protaceratherium minutum) (на переднем плане) и брахипотерия (Brachypotherium aurelianense). Автор — М. Антон.
Brachypotherium aurelianense Nouel, 1866 — вид, известный из раннего - среднего миоцена (бурдигальский - лангский ярусы) Португалии (Лиссабонский регион), Испании (автономный регион Арагон и автономное сообщество Каталония), Франции (регион Центр), Германии (земля Баден-Вюртемберг), Казахстана (Тургай) и Китая (провинция Цзянсу). Родовая принадлежность данного вида спорна. Антунес и Гинзбург (1983 г.), а также Лу с соавторами (2021 г.) полагают, что он должен быть включен в род Diaceratherium в качестве Diaceratherium aurelianense, тогда как Цердено с соавторами (1995 г.) относят его к роду Brachypotherium.
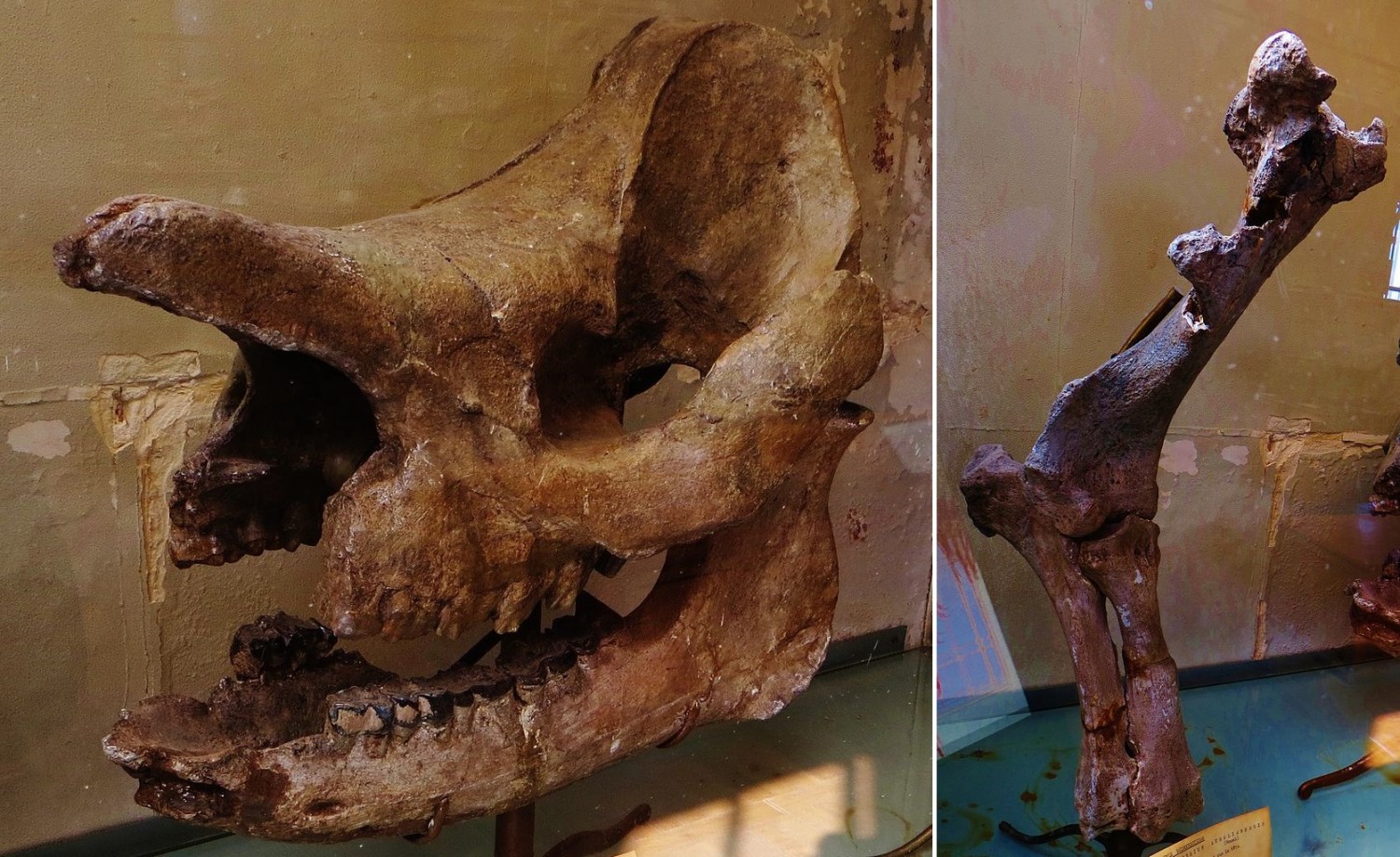
Череп с нижней челюстью и часть передней конечности брахипотерия (Brachypotherium aurelianense).
Общая длина тела составляла около 2,8 м, а высота в холке достигала 1,25 м. Череп от кончика носовых костей до затылочного гребня имеет около 45 см в длину, его скуловая ширина равняется 35 см. Носовые кости на конце утолщены и обладают характерной мозолистостью, а над глазами ближе ко лбу расположен округлый выступ со слабой шероховатостью. Это указывает на наличие двух рогов — узкого носового и меньшего лобного. Имеется отчетливое антерокроше на P3-4. Металоф на Р2-4 вдавленный, на Р4 длинный. М2 с мезостилем. Коренные зубы без язычного пояска.
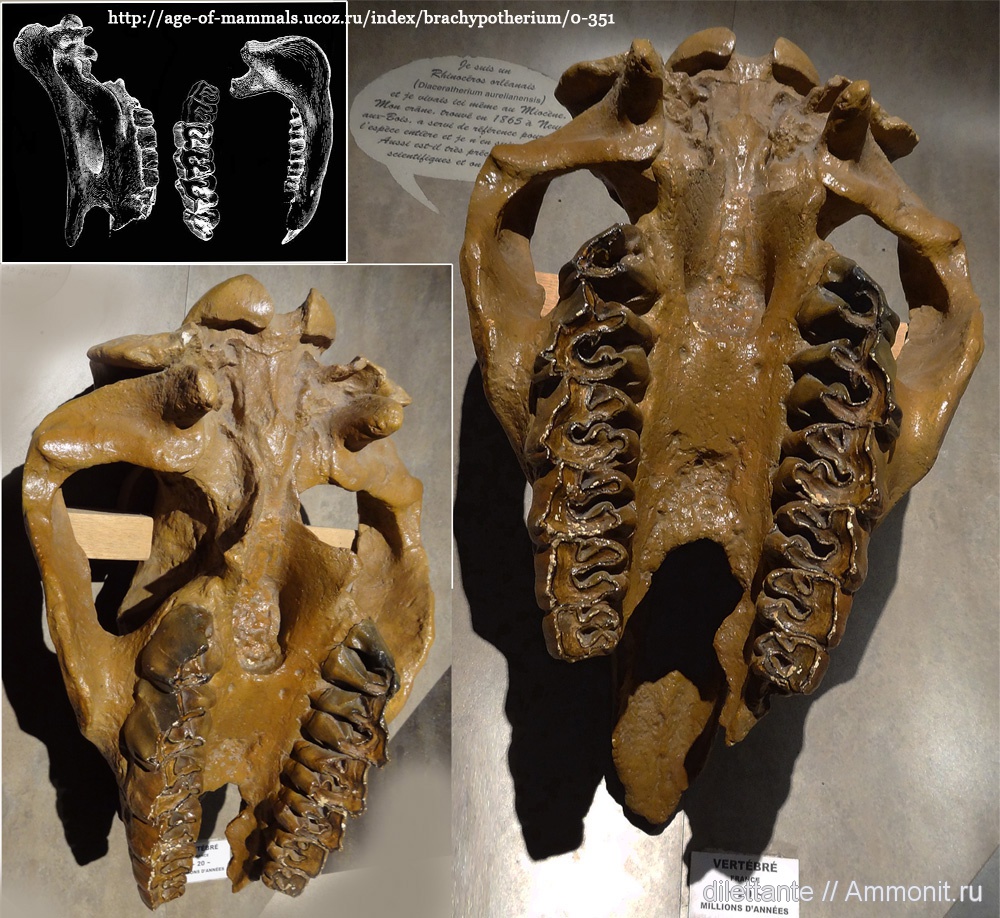
Черепа брахипотерия (Brachypotherium aurelianense).
Brachypotherium brachypus (Lartet, 1837) Lartet, 1848 — типовой вид, известный из раннего - позднего миоцена (бурдигальский - тортонский ярусы) Франции (регионы Гранд-Эст, Центр, Окситания, Аквитания, Пеи-де-ла-Луар), Германии (земли Баден-Вюртемберг, Бавария), Австрии (земля Каринтия) и Польши (Силезия). Впервые описан из позднего миоцена Сансана, Франция. Синоним: Rhinoceros brachypus Lartet, 1837 (тип).
Размеры довольно крупные: длина m1-2 составляет 70 мм, масса тела достигала 1,5 т. Антерокроше редуцировано или отсутствует. Внутренний поясок довольно отчетливый. Как и у Brachypotherium aurelianense, металоф Р4 длинный.

Нижняя челюсть брахипотерия (Brachypotherium brachypus).
Brachypotherium stehlini Roman et Viret, 1934 — сомнительный вид, известный из раннего / среднего миоцена (верхи бурдигальского / лангский ярус) Франции (местонахождения Ла-Ромэ и Беньо). Вполне вероятно, что данная форма на самом деле является подвидом и должен указываться как Brachypotherium brachypus stehlini. Метаподии относительно длиннее, чем у Brachypotherium brachypus.
Brachypotherium goldfussi Kaup, 1834 — вид, известный из позднего миоцена / раннего плиоцена (мессинский / занклский ярус) Германии (земля Баден-Вюртемберг). Описан по 2 нижним коренным зубам из Эппельсгейма. Судя по размерам найденных зубов (их общая длина составляет около 60 мм), это был довольно крупный носорог, хотя и мельче, чем Brachypotherium aurelianense. Он был последним представителем рода в Европе.
Brachypotherium snowi Fourtau, 1918 — вид, известный из раннего - среднего миоцена (бурдигальский - серравалльский ярусы) Египта (местонахождения Вади-Могара и Сива) и Ливии (Джебель-Зелтен). Кроме того, возможно, к этому виду принадлежат и неописанные останки из Мвити в Каджонге, Кения. Синоним: Aceratherium campbelli Hamilton, 1973.
Размеры крупные: длина ряда щечных зубов составляет около 270 мм, длина М1-2 — 70 мм. Череп широкий и низкий. Носовая вырезка доходит до P3. Носовые кости длинные, вероятно, с небольшим рогом или парой рогов. Передний край глазницы расположен на уровне M2. Скулы мощные и очень широкие. Верхний профиль черепа сильно вогнутый. Височные линии почти сливаются в сагиттальной гребень. Затылок округлый. Brachypotherium snowi демонстрирует некоторое сходство с европейским Prosantorhinus, но четко отличается более низким и широким черепом, более выраженной шероховатостью на носовых костях и более короткими метаподиями. Данный вид является предшественником и наиболее вероятным предком Brachypotherium lewisi.

Реконструкции внешнего облика носорогов дицерорина (Dicerorhinus leakey) (на заднем плане) и брахипотерия (Brachipotherium heinzelini). Автор — М. Антон.
Brachypotherium heinzelini Hooijer, 1963 — вид, известный из раннего - среднего миоцена (бурдигальский - серравалльский ярусы) Демократической Республики Конго, Кении и Уганды. Описан по Р4 из раннего миоцена Синда-Мохари, Синда-Онголиба, бассейн реки Семлики в ДРК. Помимо этого, фрагментарные остатки известен из среднего миоцена местонахождений Русинга, Карунгу, Буква и Напак II в Кении. В настоящее время высказываются мнения, что Brachypotherium heinzelini из Синда следует считать младшим синонимом Brachypotherium lewisi (Пикфорд с соавт., 1993), тогда как другие образцы необходимо отнести к Brachypotherium snowi. Как и у Brachypotherium аurelianense, антерокроше хорошо развито.
Brachypotherium minor Geraads et Miller, 2013 — вид, известный из раннего миоцена Кении. Описан по относительно полному черепу из отложений формации Бакате местонахождения Булук на севере страны. Некрупный представитель рода, морфологически сходный с Brachypotherium aurelianense.

Череп брахипотерия (Brachypotherium minor) KNM WS-46072 (типовой образец) из Булука, Кения: A — сверху; B — сзади; C — слева; D — небная поверхность с левыми P2-M3. Длина масштабной линейки: A-C — 20 см; D — 10 см.
Brachypotherium lewisi Hooijer, 1972 — вид, известный из позднего миоцена - раннего / ? позднего плиоцена (тортонский - занклский / ? пьяченцский ярусы) Ливии, Эфиопии и Кении. Описан по 2 неполным черепам, нижней челюсти и элементам посткраниального скелета из отложений формации Навата в Лотагаме, Кения, другие находки совершены в более ранних и поздних горизонтах Лотагама, Нгороре и Мпесиде (Кения), а также в Сахаби (Ливия) и в среднем течении реки Аваш (Эфиопия).
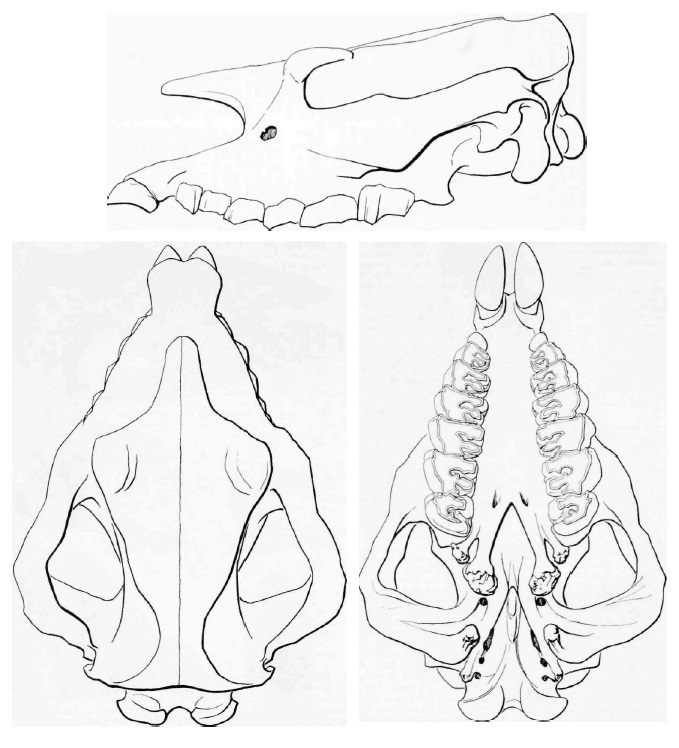
Прорисовки реконструированного черепа брахипотерия (Brachypotherium lewisi) из Кении.
Очень крупный вид и один из самых больших представителей всего семейства. Скелет массивнее, чем у других вымерших и современных африканских носорогов. Расчетная длина тела — 3,5 м, высота в холке — 1,5 м, масса — около 3 т. Кондилобазальная длина черепа превосходит 70 см, длина М1-2 составляет 90 мм, ширина неба между M3 — 120 мм. Носовая вырезка узкая, но не очень длинная (однако длиннее, чем у Brachypotherium аurelianense), ее наиболее глубокая точка расположена над Р4. Носовой рог отсутствует. Передний край глазницы расположен на уровне M2. Нижняя лопасть чешуйчатой кости (часть височной кости) проходит ниже субаурикулярного канала. Верхние резцы очень большие. Верхние щечные зубы брахиодонтные. Эктолоф уплощенный за параконом, антекроше умеренное, протокон сужен незначительно, часто присутствует внешний эмалевый поясок. Brachypotherium lewisi отличается от Brachypotherium snowi большими размерами, прямым верхним профилем черепа, V-образной хоаной, более глубокой носовой вырезкой, короткой диастемой, отсутствием i1 и меньшим i2.
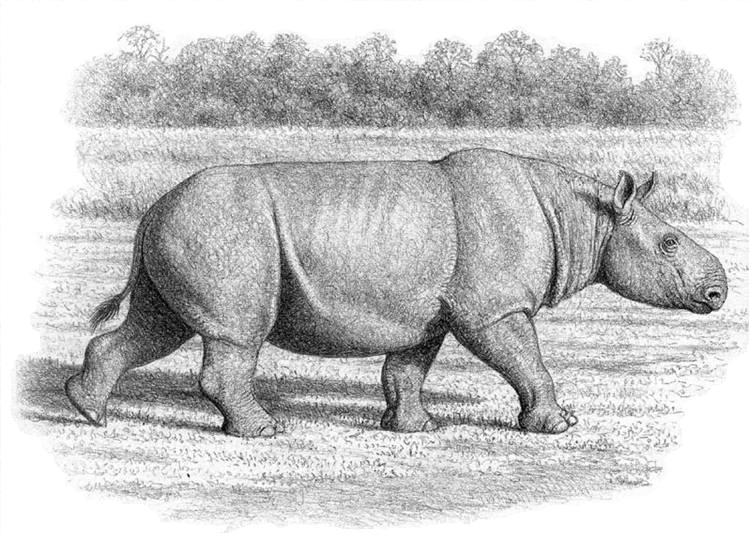
Брахипотерий (Brachypotherium lewisi) в естественном окружении. Автор — М. Антон.
Остатки, определенные лишь до рода и обозначенные Brachypotherium spp., известны из позднего олигоцена - позднего миоцена Германии (земля Баден-Вюртемберг), Чехии, Словакии, Турции, Казахстана, Таджикистана, Китая (провинции Цинхай и Шэньси), Японии (регион Канто на острове Хонсю), Мьянмы, Таиланда, Туниса, Уганды и Намибии. Некоторые из этих находок спорные, на основании других со временем могут быть выделены новые виды.

Нижняя челюсть брахипотерия (Brachypotherium sp.).
Образ жизни
Брахипотерии являлись типичным компонентом миоценовой фауны. Они обитали в саваннах и лесах с редкими полянами, где придерживались берегов водоемов. Строение кисти и стопы позволяло без особых затруднений передвигаться по топкому грунту, а значительный объем туловища придавал дополнительную плавучесть в водной среде. Подобно современному гиппопотаму (Hippopotamus amphibius), значительную часть времени брахипотерии могли проводить в воде или грязи, оставаясь довольно неуклюжими на суше. Судя по относительно низким и просто устроенным щечных зубам, в рацион этих носорогов входила как мягкая, так и жесткая растительность: листья, почки, тонкие веточки деревьев и кустарников, а также травянистые околоводные и водные растения.

Брахипотерии (Brachypotherium sp.) в естественном окружении.
Вымирание
Наиболее вероятной причиной исчезновения брахипотериев являются климатические изменения, повлекшие перестройку растительных сообществ. Потеряв привычную среду, эти носороги не смогли приспособиться к новым условиям и вымерли.
Ссылки
Brachypotherium (Paleobiology Database) перейти
Brachypotherium (Wikipedia) перейти
Литература
Основы палеонтологии. В 15 т. Т. 13: Млекопитающие / Гл. ред. Орлов Ю. А. — М.: Государственное научно-техническое издательство литературы по геологии и охране недр, 1962
Cenozoic Mammals of Africa. Edited by Werdelin L. & Sanders W. J. — Berkeley: University of California Press, 2010
Cerdeño E. Spanish Neogene rhinoceroses. Palaeontology 35 (2), 1992
Cerdeño E. Etude sur Diaceratherium aurelianense et Brachypotherium brachypus du Miocene Moyen de France. Bulletin du Museum National d'Histoire Naturelle de Paris Series 4 (15) C (1-4), 1993
Cerdeño E. Diversity and evolutionary trends of the the family Rhinocerotidae (Perissodactyla). Palaeo 141, 1998
Deng T., Downs W. Evolution of Chinese Neogene Rhinocerotidae and its response to climatic variations. Acta geologica sinica 76 (2), 2002
Deng T. Incisor fossils of Aprotodon (Perissodactyla, Rhinocerotidae) from the Early Miocene Shangzhuang Formation of the Linxia Basin in Gansu, China. Vertebrata Palasiatica, Beijing 51 (2), 2013
Evolution of Tertiary Mammals of North America. Vol. 1: Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals / Ed. by Janis C. M., Scott K. M. & Jacobs L. L. — Cambridge: Cambridge University Press, 1998
Fortelius M. Rhinocerotidae from Pasalar, middle miocene of Anatolia (Turkey). Journal of Human Evolution 19, 1990
Fukuchi A., Kawai K. Revision of Fossil Rhinoceroses from the Miocene Mizunami Group, Japan. Paleontological Research 15 (4), 2011
Geraads D., Saraç G. Rhinocerotidae from the Middle Miocene Hominoid Locality of Çandir (Turkey). Courier des Forschungsinstituts Senckenberg 240, 2003
Geraads D., Spassov N. Rhinocerotidae (Mammalia) from the Late Miocene of Bulgaria. Palaeontographica A, 287, 2009
Geraads D., Miller E. Brachypotherium minor n. sp., and other Rhinocerotidae from the Early Miocene of Buluk, Northern Kenya. Geodiversitas 35 (2), 2013
Heissig K. Brachypotherium aus dem Miozän von Südwestafrika. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historischer Geologie 11, 1971
Heissig K. Paläontologische und geologische Untersuchungen im tertiär von Pakistan. 5. Rhinocerotidae (Mamm.) aus den unteren und mittleren Siwalik-Schichten. Bayerische Akademie der Wissenschaften, mathematisch-naturwissenschaftliche Klasse, Abhandlungen 152, 1972
Heissig K. The early Vallesian vertebrates of Atzelsdorf (Late Miocene, Austria) 11. Rhinocerotidae and Chalicotheriidae (Perissodactyla). Annalen des Naturhistorischen Museums zu Wien 111 A, 2009
Hooijer D. A., Patterson B. Rhinoceroses from the pliocene of Northwestern Kenya. Bulletin of the Museum of Comparative Zoology, Harvard University 144 (1), 1972
Khan A. M., Akhtar M., Khan M. A., Shaheen A. New fossil remains of Brachypotherium perimense from the Chinji and Nagri formations of Pakistan. The Journal of Animal & Plant Sciences 22 (2), 2012
Khan A. M., Habib A., Khan M. A., Ali M., Akhtar M. New remains of Brachypotherium fatehjangense from Lower Siwalik Hills, Punjab, Pakistan. The Journal of Animal & Plant Sciences, 20 (2), 2010
Leakey M. G., Harris J. M. Lothagam: the dawn of humanity in eastern Africa. New York: Columbia University Press, 2003
Osborn H. F. Phylogeny of the rhinoceroses of Europe. Bulletin of the American Museum of Natural History 13 (19), 1900
Pilgrim G. E. The vertebrate fauna of the Gaj Series in the Bugti Hills and the Punjab. Memoirs of the Geological Survey of India (Palaeontologia Indica) New Series 4 (2), 1912
Qiu Z., Xie J. A new species of Aprotodon (Perissodactyla, Rhinocerotidae) from Lanzhou Basin, Gansu, China. Vertebrata Palasiatica 35 (4), 1997
The Evolution of Perissodactyls / Ed. by Prothero D. R. & Schoch R. M. — New York: Oxford University Press, 1989
The Miocene land mammals of Europe / Ed. by Rössner G. E. & Heissig K. — München: Pfeil, 1999
Yatkola D., Tanner L. G. Brachypotherium from the Tertiary of North America. Occasional papers of the Museum of Natural History University of Kansas 77, 1979
Zervanová J., Sabol M., Hudáčková-Hlavatá N., Holec P. Brachypotherium cf. brachypus and Lartetotherium sp. (Rhinocerotidae, Perissodactyla, Mammalia) from the Middle Miocene Dúbravka-Pole site (western Slovakia). Acta Geologica Slovaca 5 (1), 2013
Zin-Maung-Maung-Thein, Taru H., Takai M., Fukuchi A. A Rhinocerotid (Mammalia, Perissodactyla) from the Late Miocene Oiso Formation, Kanagawa Prefecture, Japan. Paleontological Research, 13 (2), 2009