Бронтотериевые — Brontotheriidae
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Клада Скротиферы (Scrotifera)
Клада Феревунгуляты (Fereuungulata)
Клада Эвунгуляты (Euungulata)
Клада Мезаксонии (Mesaxonia)
Отряд Непарнопалые (Perissodactyla)
Подотряд †Лофодонтообразные (Lophodontomorpha)
Инфраотряд †Эвпериссодактили (Euperissodactyla)
Клада Лошадеобразные (Hippomorpha)
Надсемейство †Бронтотериоиды (Brontotherioidea)
Семейство †Бронтотериевые (Brontotheriidae)
Бронтотериевые, или бронтотерии (Brontotheriidae Marsh, 1873), — семейство надсемейства бронтотериоидов (Brontotherioidea (Marsh, 1873)), происходящее из раннего - позднего эоцена Северной Америки, раннего эоцена - раннего олигоцена Азии и среднего - позднего эоцена Европы. Установлены 3 подсемейства и порядка 50 родов.
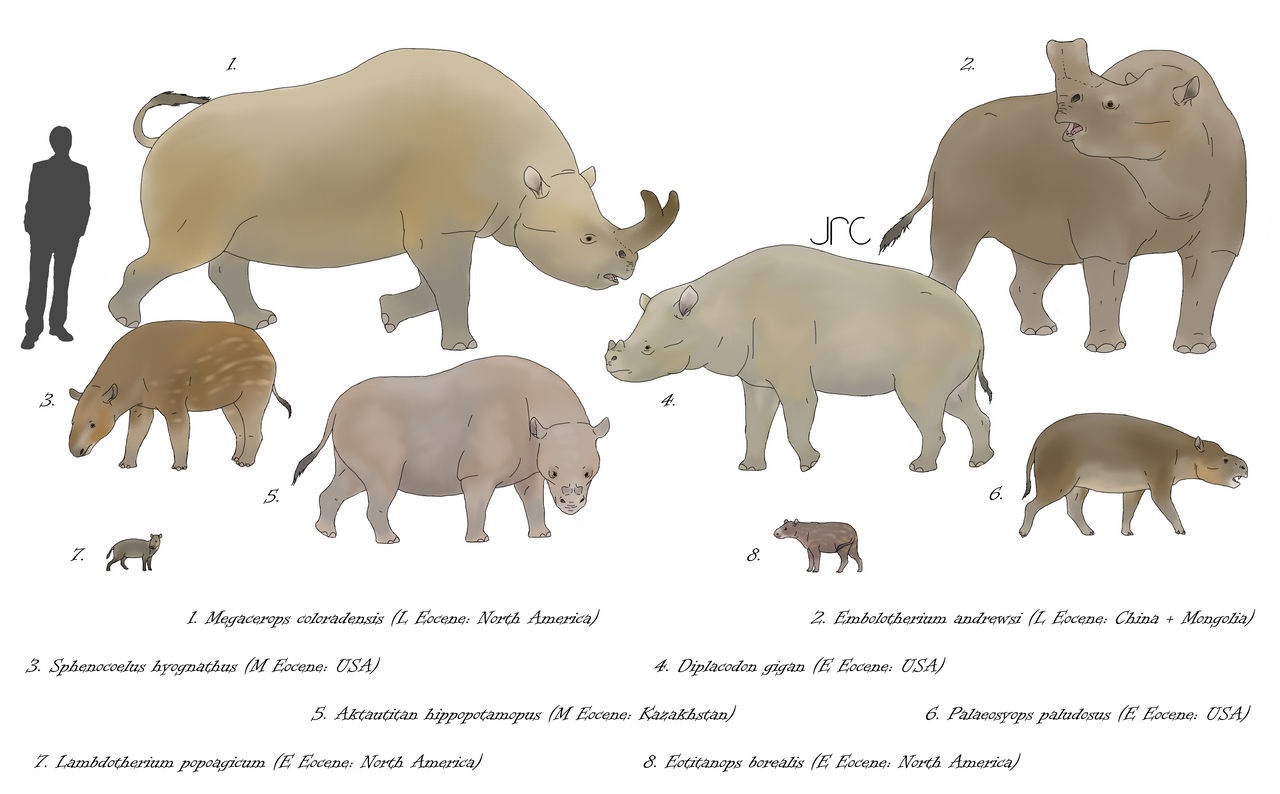
Разнообразие бронтотериоидов (Brontotherioidea). Автор — artbyjrc.
Синонимы: Titanotheriidae; Menodontidae Cope, 1881; Brontopinae Osborn, 1914.
История изучения
Ныне невалидное родовое наименование Brontotherium, давшее название всей группе, происходит от древнегреческих слов βροντή — "гром" и θηρίον — "зверь", означая в переводе "громовой зверь". Это название связано с фольклором североамериканских индейцев сиу. Они считали, что животные, кости которых обнажили дожди, производят гром, бегая над облаками, поэтому называли их "громовыми конями". Многие из скелетов, найденные сиу, принадлежали стадам бронтотериев, погибшим в результате извержений в Скалистых горах, которые в то время были вулканически активными.

Извлечение остатков мегацеропса (Megacerops coloradensis) в ходе раскопок.
В США бронтотериев начали изучать в середине XIX в. Они были одними из первых ископаемых позвоночных, известных с запада страны. Впервые окаменелости бронтотериев изучил Лейди, но доступный ему материал был довольно фрагментарным и не позволял в полной мере представить, что это были за животные. Коп и Марш были первыми, кто исследовал черепа и скелеты бронтотериев. Марш первым отнес их к непарнопалым и дал название семейству. Однако в это семейство он включил только поздние рогатые формы из формации Уайт-Ривер. Ранние примитивные формы были отнесены Маршем к Limnohyidae, однако он отмечал филогенетическую связь между ранними безрогими и поздними рогатыми формами. Примерно в то же время было выделено несколько синонимичных семейств, таких как Titanotheriidae и Menodontidae. Огромный вклад в изучение бронтотериев внес Генри Фэрфилд Осборн, написавший обширную работу об этих животных. Осборн выделял большое количество подсемейств, родов и видов бронтотериев. Однако впоследствии его классификация была упразднена и многие таксоны были признаны синонимами, а различия между образцами сейчас принято объяснять половыми и возрастными различиями.
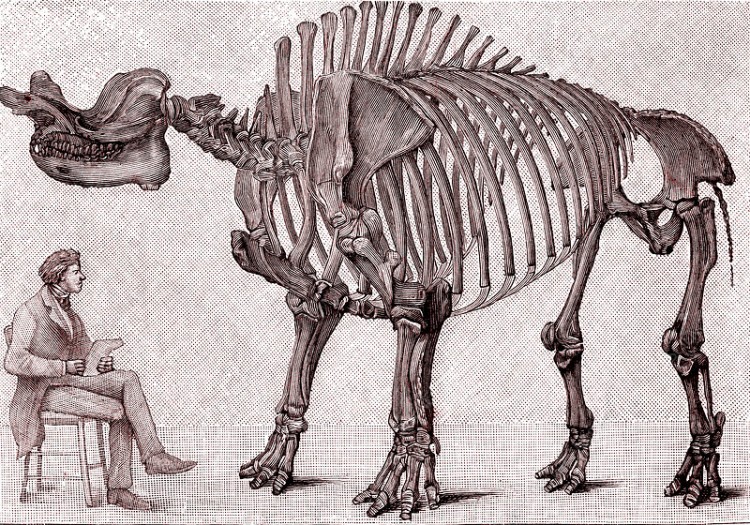
Прорисовка смонтированного скелета мегацеропса (Megacerops coloradensis, = Brontops robustus) из позднего эоцена штата Южная Дакота, США; 1891 г.
Американские бронтотерии были уже довольно хорошо известны к началу ХХ в. Первые азиатские представители семейства были открыты только в 1912 г. в отложениях Пондаунгской формации на территории современной Мьянмы. В 1920-30-х гг. экспедиции Американского музея естественной истории обнаружили большое количество остатков бронтотериев в Центральной и Юго-Восточной Азии, не менее разнообразных, чем их американские родственники. Осборн пытался отнести азиатских бронтотериев к североамериканским родам. После его смерти Гренжер и Грегори сочли это ошибкой, но они все же выделили много новых родов и видов, иногда основанные на деформированных остатках и возрастных стадиях одного вида. В дальнейшем многие палеонтологи, прежде всего китайские и российские, продолжили изучение азиатских бронтотериев и описали новые их таксоны.

Работа над пластической портретной реконструкцией мегацеропса (Megacerops coloradensis).
Происхождение и родственные связи
Филогенетическое положение бронтотериев среди непарнопалых окончательно невыяснено. Традиционно их сближают с лошадьми (Equidae) в пределах группы лошадеобразных (Hippomorpha), куда также включают палеотериев (Palaeotheriidae) и иногда халикотериев (Chalicotheriidae). Порой же их относят к носорогообразным (Ceratomorpha), куда входят носороги (Rhinoceratoidea) и тапиры (Tapiroidea). Наконец, некоторые исследователи считают, что бронтотерии не входят ни в одну из указанных групп и являются сестринским таксоном для остальных непарнопалых. Их внешнее подобие носорогам в данном случае объясняется параллельной эволюцией в сходных экологических нишах.
На сегодняшний день не представляется возможным точно сказать, откуда происходят бронтотерии. Сейчас преобладает версия североамериканского их происхождения, поскольку наиболее древние надежно определенные представители семейства (Eotitanops и др.), а также сестринский таксон и возможно предковый для бронтотериев род Lambdotherium известны из Северной Америки. Однако в Азии также обнаружены остатки ранних бронтотериев и лямбдотериеподобных копытных. Но большинство этих остатков очень фрагментарны и определены с недостаточной степенью точности. Поэтому нужно ждать новых находок, которые помогут более точно определить родину бронтотериевых.
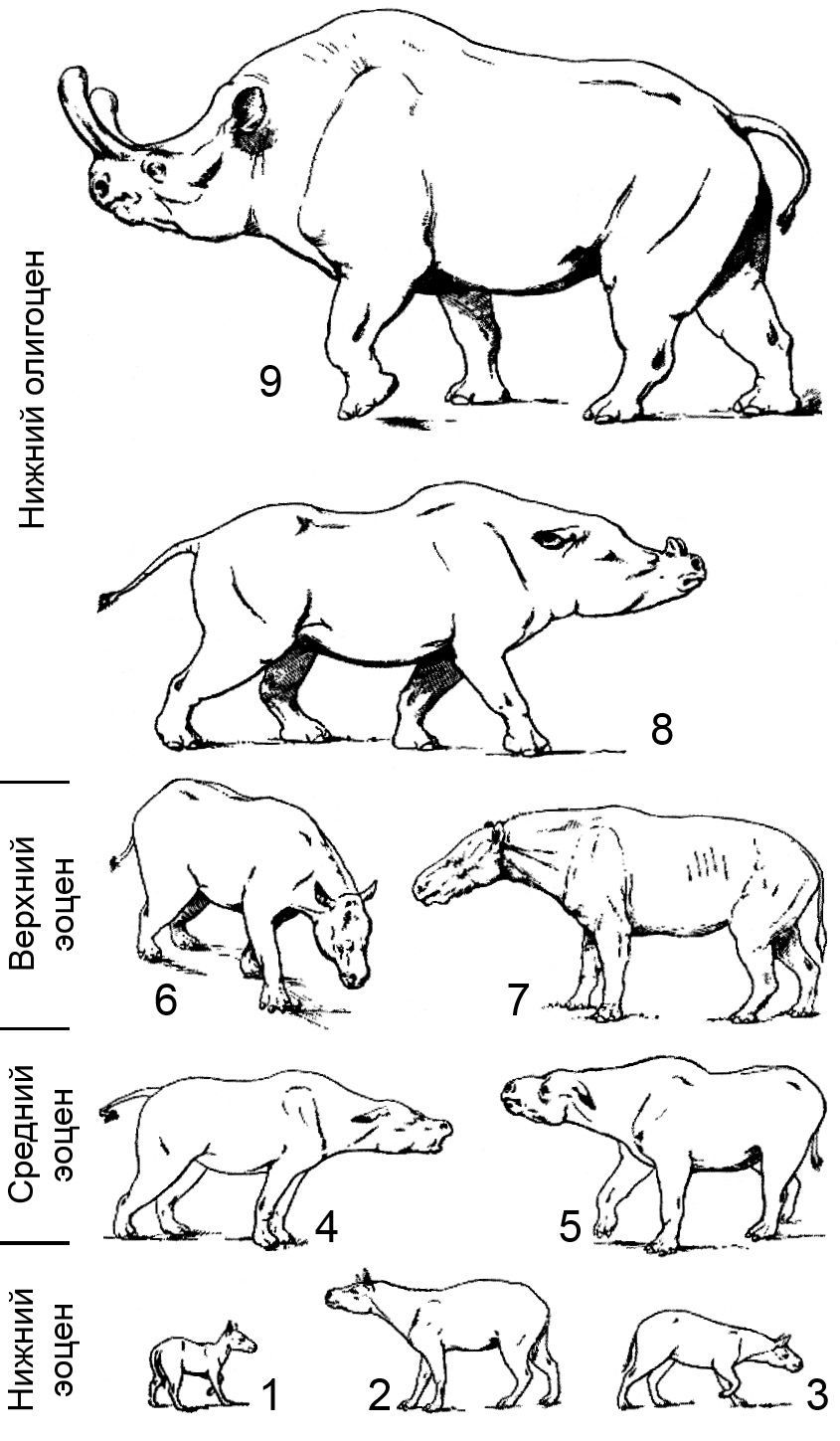
Этапы эволюции североамериканских бронтотериев (по Осборну, 1929): 1 — Lambdotherium popoagicum; 2 — Eotitanops borealis; 3 — Eotitanops minimus; 4 — Mesatirhinus junius; 5 — Palaeosyops leidyi; 6 — Telmatherium validus; 7 — Dolichorhinus hyognathus; 8 — "Brontotherium leidyi"; 9 — Megacerops coloradensis.
Распространение
Большая часть родов бронтотериев происходит из Центральной Азии и запада Северной Америки. Отдельные представители семейства известны из Юго-Восточной Европы, дальнего востока России, Пакистана, Юго-Восточной Азии, Японии, юго-востока США и севера Канады. Таким образом, бронтотерии имели широкое распространение в Голарктике за исключением Западной Европы.

Старинная схема ареала бронтотериев. Черным цветом закрашены основные области обнаружения ископаемых остатков — запад США, пустыня Гоби в Монголии, Мьянма и юго-восточная Европа. Штриховкой отмечены вероятные области распространения.
Строение и функции
Размеры бронтотериев варьировали от небольших до очень крупных: их общая длина составляла 60-500 см, высота в холке — 30-240 см, вес оценивается в 10-4500 кг. С течением времени размеры тела прогрессивно увеличивались.
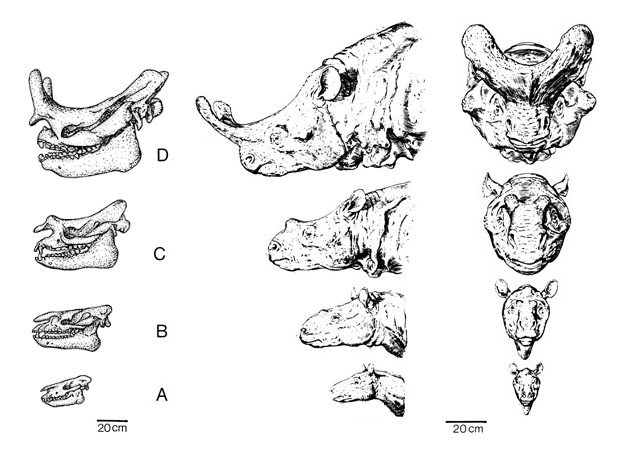
Развитие рогообразных выступов на морде у бронтотериев: A — Eotitanops (ранний эоцен, 55 млн. лет назад); B — Manteoceras (начало среднего эоцена, около 50 млн. лет назад); C — Protitanotherium (конец среднего эоцена, 45 млн. лет назад); D — Megacerops (поздний эоцен, 35 млн. лет назад).
Череп с укороченным лицевым и удлиненным мозговым отделами. Как и у носорогов, происходило постепенное укорочение предглазничной части черепа. Черепной профиль вогнутый, в форме седла, затылок часто высоко поднят. Скуловые дуги могут быть широко расставлены в стороны. Полость головного мозга невелика.
К концу среднего эоцена в нескольких линиях бронтотериев наверху морды развились костные структуры наподобие рогов, иногда достигающие внушительных размеров. У некоторых родов (Megacerops и др.) носовые и лобные кости несут одну пару рогообразных выростов, достигающие различной длины. У других (Embolotherium и др.) имеется единственный большой вырост, представляющий собой сильно увеличенные и преобразованные носовые кости, которые поднимаются вверх и лопатообразно расширяются, перекрываясь сзади лобными костями. Внутри рогообразных выростов имеются полости, заполненные губчатой костной тканью. Следует отметить, что лобно-носовые выросты бронтотериев можно назвать рогами лишь условно, поскольку они имеют мало общего с рогами современных копытных. Согласно одной точке зрения, выросты были покрыты кожей и слабо питались кровеносными сосудами, подобно оссиконам современых жирафов. По другим представлениям, у части родов рогообразные выросты покрывал плотный слой рогового вещества, что делало их гораздо более прочным.

Череп с нижней челюстью мегацеропса (Megacerops coloradensis).

Череп с нижней челюстью эмболотерия (Embolotherium andrewsi).
Зубная формула: I3-0/i3-0, C 1/c1, P4-3/p4-3, M 3/m3. Резцы у поздних форм уменьшены в числе или полностью исчезают. Клыки мелкие или средней величины. Щечные зубы брахиодонтные (низкокоронковые), широкие, плоские, бугорчато-лунчатые, с замкнутыми корнями. Предкоренные никогда полностью не морляризуются. Верхние коренные с эктолофом W-образной формы, изолированными коническим прото- и гипоконом и иногда с небольшим протоконулем; соотоношение и величина прото- и гипокона различно. Жевательная поверхность нижних коренных состоит из двух V-образных полулуний, соединенных концами в середине зуба, без ясных метаконида и метастилида.
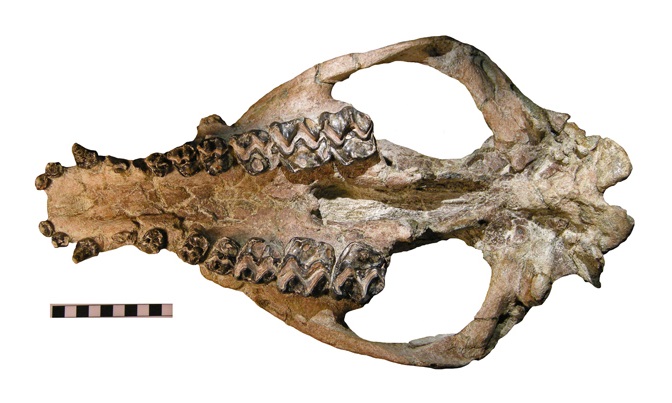
Череп тельматерия (Telmatherium validus), снизу.
Посткраниальный скелет у наиболее ранних форм легко построенный, но очень скоро становится весьма массивным. У поздних представителей группы на передних грудных позвонках развились длинные остистые отростки, к которым крепились мышцы и связки, поддерживающие голову. Конечности относительно короткие, прогрессивно укорачиваются и делаются массивнее. Кисть четырехпалая, стопа трехпалая; все конечности пальцеходящие. Средний палец лишь немногим массивнее других. Дистальные фаланги мелкие, копытообразные. Относительный консерватизм конечностей и четырехпалая кисть связан с массивным строением животных.

Поперечные срезы носовых костей различных бронтотериев.
Образ жизни
Пока нельзя уверенно утверждать, какой образ жизни вели бронтотерии. Вероятно, первые примитивные представители группы обитали в лесах, где их мелкие размеры помогали им скрываться среди растительности. Однако эволюция бронтотериев была направлена на быстрое увеличение размеров тела. Возможно, это связано с переходом к жизни в открытых ландшафтах. Но сухие степи врядли подходили для бронтотериев, так как их низкокоронковые зубы не были приспособлены для постоянного питания жесткой травянистой растительностью. Скорее всего, эти копытные тяготели к открытым пространствам, которые чередовались с группами деревьев и кустарников и среди которых имелись водоемы.
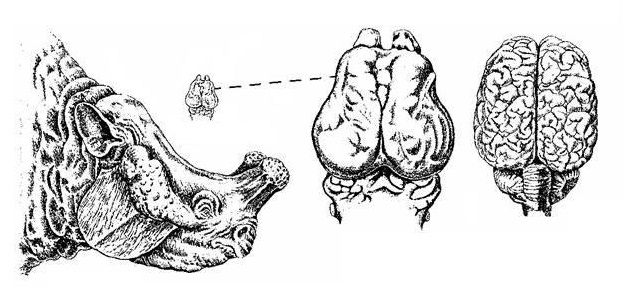
Сравнение головного мозга мегацеропса (Megacerops coloradensis) (слева) и домашней свиньи (Sus scrofa) (справа).
Ранее полагали, что некоторые бронтотерии вели полуводный образ жизни и добывали корм в воде. При этом дыхание облегчалось носздрями, расположенными на конце костного выроста черепа. Зубы бронтотериев считались слабыми, не приспособленными к питанию твердой пищей, из-за недостаточной гипсодонтности. Однако, учитывая крупные размеры большинства видов, можно предположить, что они были не очень разборчивыми едоками. Вероятнее всего, бронтотеривеые питались листьями, молодыми побегами и, возможно, мягкими плодами кустарников и невысоких деревьев.

Следовые отпечатки бронтотерия из позднеэоценовых отложений формации Грин-Ривер, округ Севи, штат Юта, США.
Полагают, что бронтотерии жили стадами и им была свойственна полигамия. Этим может объясняться ярко выраженный половой диморфизм, выраженный в размерах тела, строении черепа и особенно рогообразных выростов на морде. Судя по всему, самцы обладали более крупными выростами, имеющими демонстрационные и опознавательные функции. Высказано предположение, что бронтотерии могли использовать свои костные выросты для лобовых столкновений подобно современным баранам. Изменение их формы связывают с постепенным приспособлением к возрастанию размеров всего организма. Роды, у которых рогообразные выросты достигали значительной величины и отчетливо раздваивались (например, Megacerops), могли сцепляться ими подобно тому, как сегодня сцепляются рогами олени и некоторые полорогие. Другие представители семейства могли использовать свои выросты для нанесения ударов по телу противника. Помимо внутривидовых стычек, "рога" моги использоваться для отражения атак хищников.

Пара тельматериев (Telmatherium validus) и долихорин (Dolichorhinus hyognathus) у водопоя.
Вымирание
Бронтотерии доминировали среди непарнопалых на протяжении всего эоцена. Начав свою эволюцию в начале эоцена с неспециализированных животных размером с овцу, к концу этой эпохи они достигли величины очень крупных носорогов. В этом отношении бронтотерии значительно опередили других непарнопалых. Наряду с общим укрупнением тела происходило необычное увеличение размеров костных выростов на передней части черепа. Однако, несмотря на максимальное увеличение в размерах и крайнюю специализацию в строении черепа и зубов, к началу олигоцена бронтотерии переживают сильный упадок. Область их распространения заметно сократилась. В Северной Америке бронтотерии исчезают еще до конца эоцена. В Азии они продержались немного дольше, но и там позже раннего олигоцена уже неизвестны.

Мегацеропсы (Megacerops coloradensis, = Brontotherium platyceras) в естественном окружении. Автор — Ч. Найт.
Точные причины вымирания семейства неизвестны. Возможно, одной из причин послужило характерное строение зубов этих животных. Оно было таковым, что зубы не могли стать гипсодонтными или лофодонтными, что было очень невыгодно на фоне меняющегося климата, а всед за ним и растительного покрова. По-видимому, растительность стала более жесткой и требовала более совершенного строения зубов для ее пережевывания. Некоторые невыгодные признаки наблюдались и в строении конечностей. Кроме того, бронтотерии обладали весьма примитивным мозгом, очень маленьким для такого массивного тела. В это время стали более многочисленными и разнообразными другие представители непарнопалых, в первую очередь носороги. Эти животные имели более развитый мозг, были значительно подвижнее и претендовали на сходные с бронтотериями экологические ниши. Бронтотерии не смогли конкурировать с ними. В результате — видимо, по ряду причин — группа полностью исчезла, не дожив до среднего олигоцена.

Портретная реконструкция самца мегацеропса (Megacerops coloradensis).
Состав
Генри Ферфилд Осборн выделял в семейства бронтотериевых 11 подсемейств. Однако в дальнейшем их классификация неоднократно пересматривалась, и количество подсемейств уменьшалось. То же самое происходило и с таксонами более низкого ранга. Как уже говорилось, многие роды и виды были впоследствии объединены, а различия в строении черепов признаны половыми и возрастными различиями, а также деформацией.
На сегодняшний день семейство имеет следующий состав:
род †Данджангии (Danjiangia Wang, 1995);
род †Балохтитанопсы (Balochititanops Missiaen, Gunnell et Gingerich, 2011);
род †Мулькраджанопсы (Mulkrajanops Kumar et Sahni, 1985);
род †Пакотитанопсы (Pakotitanops West, 1980);
род †Ксилотитаны (Xylotitan Mihlbachler et Samuels, 2016);
подсемейство †Эотитанопины (Eotitanopinae Osborn, 1914);
род †Эотитанопсы (Eotitanops Osborn, 1907);
подсемейство †Палеосиопины (Palaeosyopinae Steinmann et Döderlein, 1890);
род †Палеосиопсы (Palaeosyops Leidy, 1870);
подсемейство †Бронтотериины (Brontotheriinae (Marsh, 1873));
род †Бунобронтопсы (Bunobrontops Holroyd et Ciochon, 2000);
род †Мезатирины (Mesatirhinus Osborn, 1908);
род †Долихорины (Dolichorhinus Hatcher, 1895);
род †Сфеноцели (Sphenocoelus Osborn, 1895);
род †Десматотитаны (Desmatotitan Granger et Gregory, 1943);
род †Гиотитаны (Hyotitan Granger et Gregory, 1943);
род †Акротитаны (Acrotitan Ye, 1983);
триба †Бронтотериини (Brontotheriini Marsh, 1873);
род †Микротитаны (Microtitan Granger et Gregory, 1943);
род †Стенодекты (Sthenodectes Gregory, 1912);
род †Эпимантеоцеры (Epimanteoceras Granger et Gregory, 1943);
род †Нанотитанопсы (Nanotitanops Qi et Beard, 1998);
род †Протитаны (Protitan Granger et Gregory, 1943);
род †Протитанотерии (Protitanotherium Hatcher, 1895);
род †Ринотитаны (Rhinotitan Granger et Gregory, 1943);
род †Cиватитанопсы (Sivatitanops Pilgrim, 1925);
подтриба †Радинорина (Rhadinorhinina Osborn, 1929);
род †Фоссендорины (Fossendorhinus Mihlbachler, 2008);
род †Метарины (Metarhinus Osborn, 1908);
подтриба †Тельматериина (Telmatheriina Osborn, 1914);
род †Тельматерии (Telmatherium Marsh, 1872);
род †Метательматерии (Metatelmatherium Granger et Gregory, 1938);
род †Виккии (Wickia Mihlbachler, 2008);
род †Цюйфутитаны (Qufutitan Wang et Wang, 1997);
подтриба †Эмболотериина (Embolotheriina (Osborn, 1929));
род †Поллиосборнии (Pollyosbornia Mihlbachler, 2008);
род †Гнатотитаны (Gnathotitan Granger et Gregory, 1943);
род †Актаутитаны (Aktautitan Mihlbachler, Lucas, Emry et Bayshashov, 2004);
род †Пигмееетитаны (Pygmaetitan Miao, 1982);
род †Брахидиастематерии (Brachydiastematherium Böckh et Matyasovski, 1876);
род †Метатитаны (Metatitan Granger et Gregory, 1943);
род †Маобронтопсы (Maobrontops Averianov, Danilov, Chen et Jin, 2018);
род †Назамплюсы (Nasamplus Mihlbachler, 2008);
род †Протэмболотерии (Protembolotherium Tang, You, Xu, Qiu et Hu, 1974);
род †Эмболотерии (Embolotherium Osborn, 1929);
подтриба †Бронтотериина (Brontotheriina (Marsh, 1873));
род †Диплакодоны (Diplacodon Marsh, 1875);
род †Псевдодиплакодоны (Pseudodiplacodon Mader, 2000);
род †Пахититаны (Pachytitan Granger et Gregory, 1943);
род †Парвикорны (Parvicornus Mihlbachler et Deméré, 2009);
род †Парабронтопсы (Parabrontops Granger et Gregory, 1943);
род †Эвбронтотерии (Eubrontotherium Mihlbachler, 2007);
род †Протитанопсы (Protitanops Stock, 1936);
род †Нотиотитанопсы (Notiotitanops Gazin et Sullivan, 1942);
род †Дианотитаны (Dianotitan Chow, Chang et Ting, 1974);
род †Дюшенеоды (Duchesneodus Lucas et Schoch, 1982);
род †Мегацеропсы (Megacerops Leidy, 1870).
Ссылки
Бронтотериевые (Wikipedia) перейти
Brontotheriidae (Paleobiology Database) перейти
Литература
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 3 — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 13: Млекопитающие / Гл. ред. Орлов Ю. А. — М.: Государственное научно-техническое издательство литературы по геологии и охране недр, 1962
Mihlbachler M. C. Phylogenetic Systematics of the Brontotheriidae (Mammalia, Perissodactyla) (Ph.D. thesis). — Columbia University, 2004
Mihlbachler M. C. Species taxonomy, phylogeny, and biogeography of the Brontotheriidae (Mammalia: Perissodactyla). Bulletin of the American Museum of Natural History 311, 2008
Rose K. D. The Beginning of the Age of Mammals. — Oxford: Johns Hopkins University Press, 2006