Циногнаты — Cynognathus
Класс †Синапсиды (Synapsida)
Подкласс †Терапсиды (Therapsida)
Надотряд †Терозавры (Therosauria)
Отряд †Цинодонты (Cynodontia)
Клада †Эпицинодонты (Epicynodontia)
Инфраотряд †Эвцинодонты (Eucynodontia)
Клада †Циногнаты (Cynognathia)
Семейство †Циногнатовые (Cynognathidae)
Род †Циногнаты (Cynognathus)
Вид †Cynognathus crateronotus
Циногнаты (Cynognathus Seeley, 1895) — монотипический род монотипического семейства циногнатовых (Cynognathidae Watson, 1917), происходящий из раннего - среднего триаса Африки, раннего / среднего триаса Антарктики и Азии, а также позднего триаса Южной Америки.
Пара отдыхающих циногнатов (Cynognathus crateronotus). Автор — Р. Евсеев.
Синонимы: Nythosaurus Owen, 1876 (частью); Karoomys Broom, 1903; Karroomys Broom, 1903; Lycognathus Broom, 1913; Lycaenognathus Broom, 1915; Lycochampsa Broom, 1915; Cynidiognathus Haughton, 1922; Lycaenognathus Broom, 1925; Cynariognathus Broom, 1931 (частью); Cynogomphius Broom, 1932; Cistecynodon Brick et Kitching, 1953. Karoomys и Cistecynodon известны только по остаткам молодых особей.
Род находится в самом основании клады Cynognathia. Его название происходит от древнегреческого словосочетания κύνεος γνάθος и переводится как "собачья челюсть", или "клыкастая челюсть".
Единственный установленный вид — Cynognathus crateronotus Seeley, 1895 (тип). Он известен из раннего - среднего триаса (зона Cynognathus, середина оленекского - верхи анизийского яруса) ЮАР (Восточная Капская провинция), Лесото и Замбии (Восточная провинция), среднего триаса (анизийский ярус) Танзании (область Рувума), раннего / среднего триаса Китая, а также позднего триаса (формация Пуэсто-Вьехо, карнийский ярус) Аргентины (провинция Мендоса). Синонимы: Cynognathus platyceps Seeley, 1895; Cynognathus beeryi Seeley, 1895; Karoomys browni Broom, 1903; Nythosaurus browni Broom, 1912; Lycognathus ferox Broom, 1913; Lycochampsa ferox (Broom, 1913); Cynognathus ferox (Broom, 1913); Cynidiognathus longiceps Haughton, 1922; Cynognathus longiceps (Haughton, 1922); Cynidiognathus broomi Haughton, 1922; Cynognathus broomi (Haughton, 1922); Lycaenognathus platyceps (Seeley, 1895) Broom, 1925; Lycaenognathus kannemeyeri Broom, 1931; Cynognathus kannemeyeri (Broom, 1931); Cynogomphius berryi (Seeley, 1895) Broom, 1932; Cynidiognathus merenskyi Broili et Schröder, 1935; Cynognathus merenskyi (Broili et Schröder, 1935); Cynariognathus seeleyi Broili et Schröder, 1936; Cynognathus seeleyi (Broili et Schröder, 1936); Cistecynodon parvus Brink et Kitching, 1953; Cynognathus minor Bonaparte, 1969.
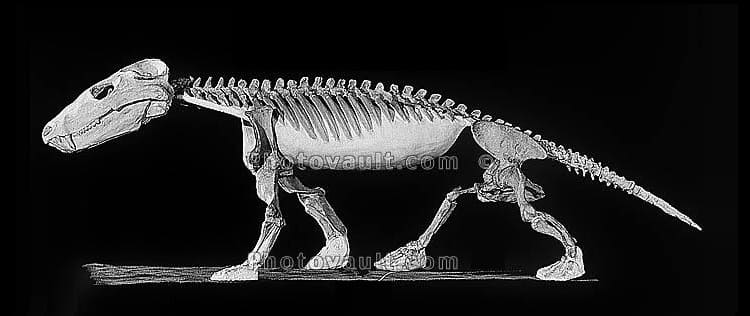
Реконструированный скелет циногната (Cynognathus crateronotus).
Остатки, определенные лишь до рода и обозначенные Cynognathus spp., известны из раннего / среднего триаса (формация Фремоу, верхи оленекского / анизийский ярус) Антарктиды (Трансантарктические горы), среднего триаса (середина - ? верхи зоны Cynognathus, низы - середина / ? верхи анизийского яруса) Намибии (область Очосондьюпа) и среднего триаса (верхи зоны Cynognathus, верхи анизийского яруса) ЮАР (Восточная Капская провинция).
Обнаружено большое количество ископаемых остатков, что позволяет реконструировать практически весь скелет. Род является диагностическим для одноименной биостратиграфической зоны триасовых отложений южноафриканской серии Карру.
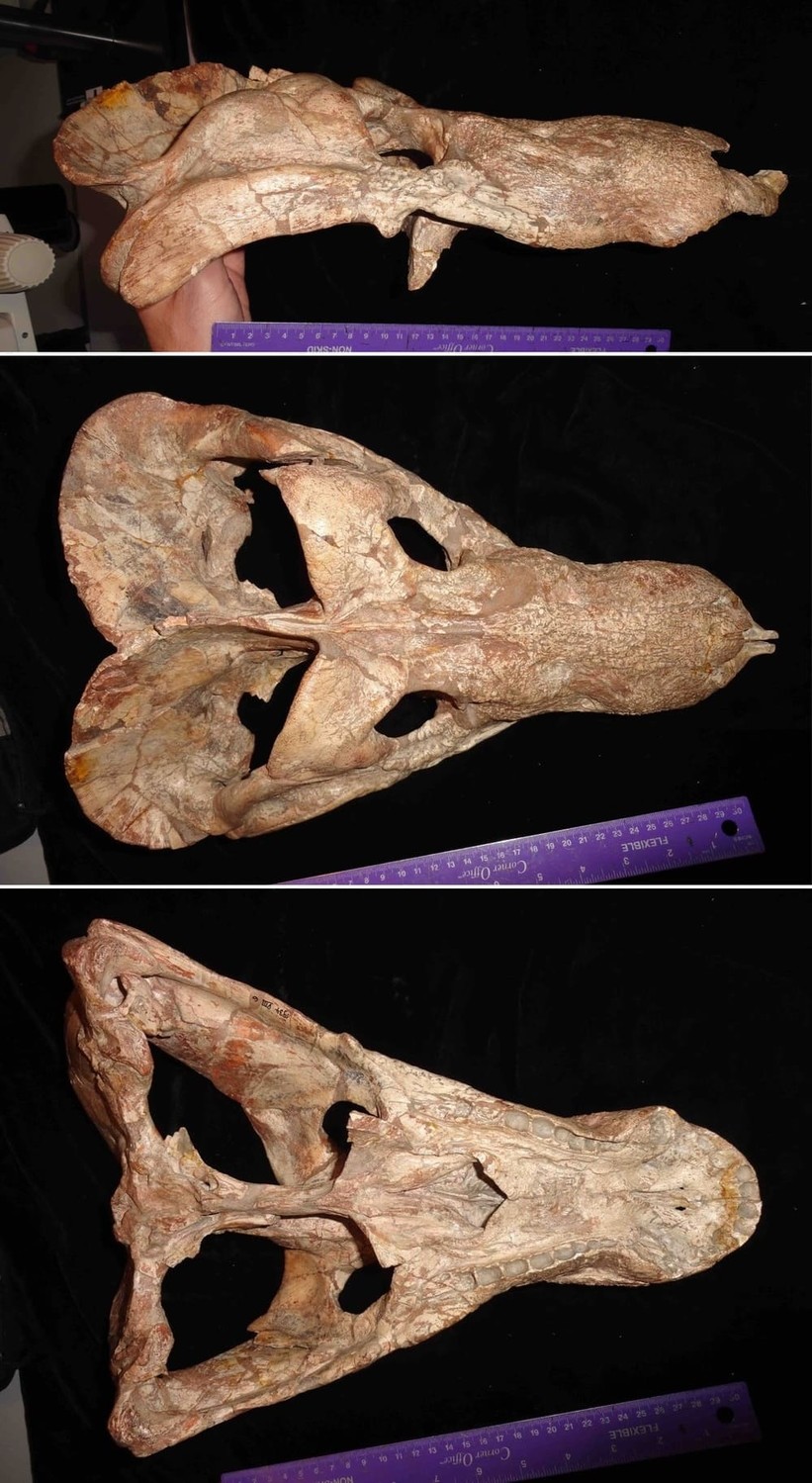
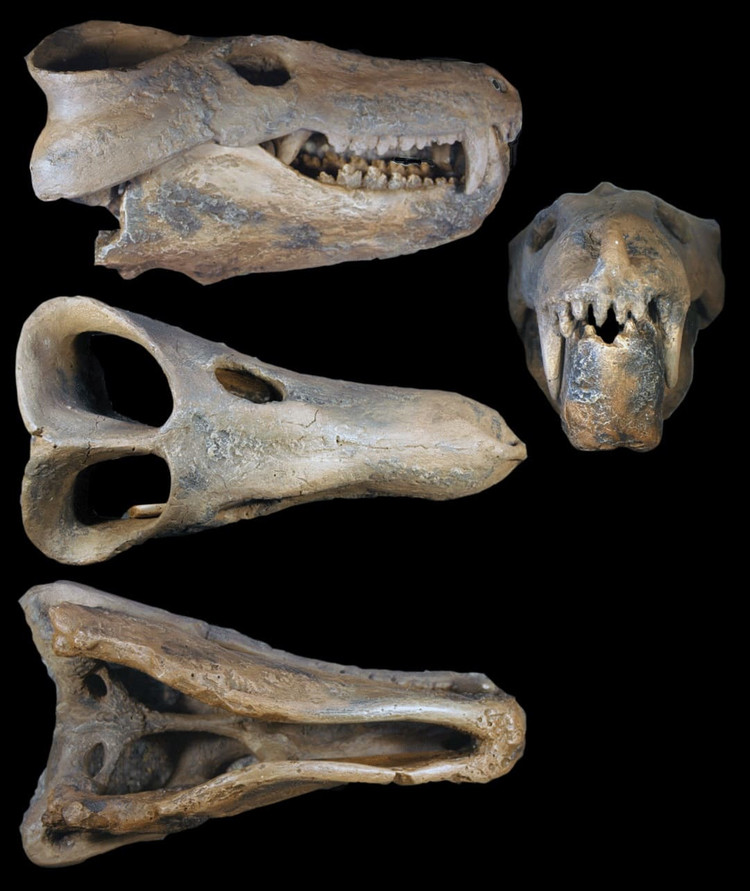
Череп с нижней челюстью циногната (
Cynognathus crateronotus).

Череп с нижней челюстью циногната (Cynognathus crateronotus) из раннего триаса ЮАР.
Череп циногната (Cynognathus crateronotus, = Cynidiognathus merenskyi) из отложений формации Бюргерсдорп, ЮАР.
Размеры варьируют от средних до крупных: череп длиной 25-40 см, общая длина составляет 1,2-2,2 м.
Череп большой относительно остального тела. Его предглазничный отдел вытянутый, сужающийся позади крупных клыков. Глазница небольшая. Заглазничная дуга широкая. Височное окно очень велико и подымается до теменной кости, заглазничная и чешуйчатая кости уже не соприкасаются друг с другом над этим окном. Окологлазничный ряд костей редуцировался; предлобная и слезная кости малы, а заднелобная исчезла. Скуловая дуга массивная и широко выступает наружу; ее верхний край расположен много ниже сагиттального гребня. Сзади скуловая дуга широко соединяется с мозговой коробкой. Аддукторная камера расширена наружу и назад; окружающие ее кости шире и толще, чем у
Thrinaxodon, что свидетельствует об очень сильном укусе. Камера не тянется так далеко вперед, значит, челюсти раскрывались шире, позволяя схватить более крупную добычу.
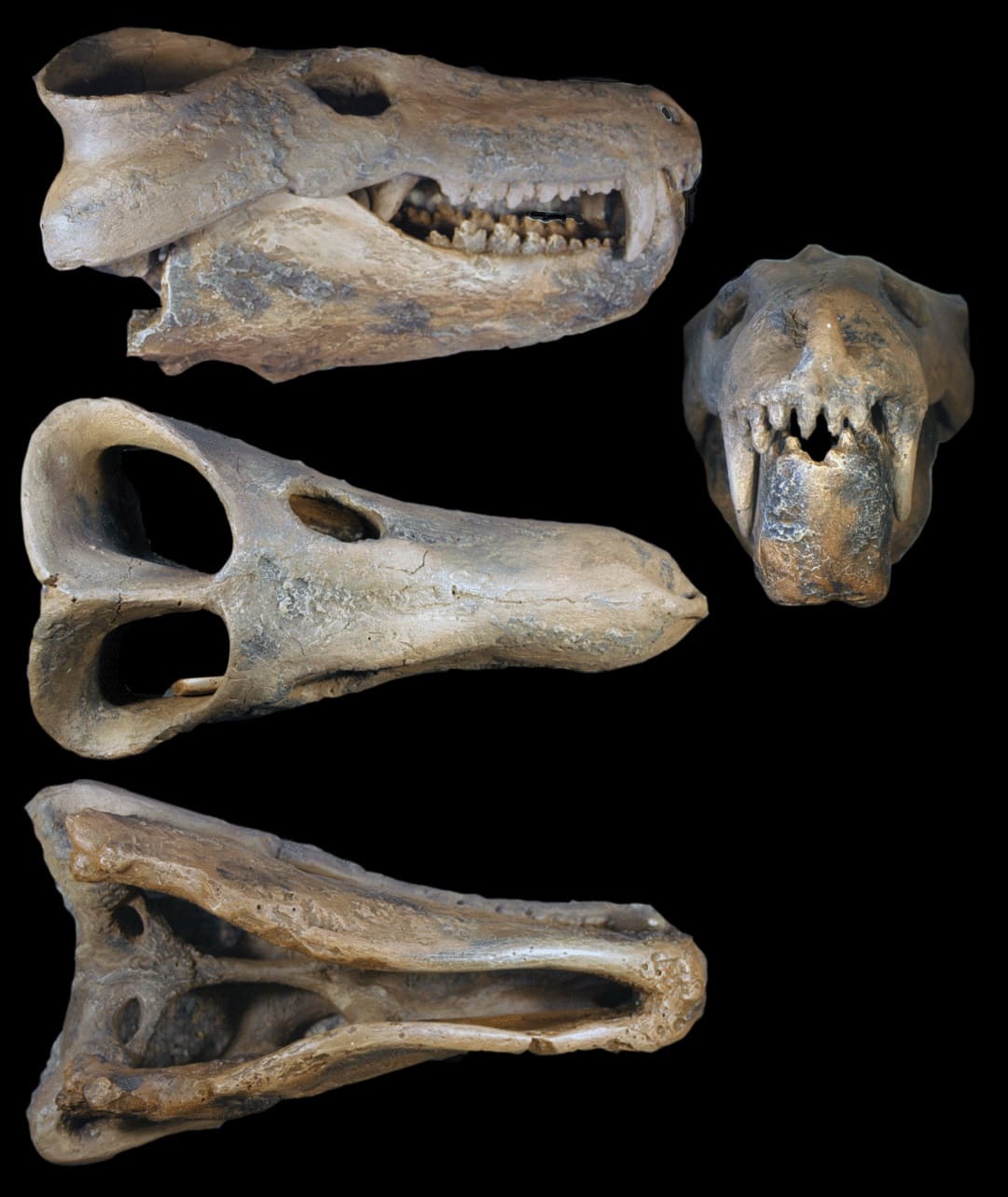
Череп с нижней челюстью циногната (
Cynognathus crateronotus).
Предтеменная кость отсутствует. Пинеальное отверстие очень мало или отсутствует. Сагиттальный гребень хорошо развит. Квадратноскуловая кость мала и тесно прирастает ко столь же редуцированной квадратной; обе эти кости довольно слабо соединены с остальным черепом. На затылочной поверхности чешуйчатой кости имеется отчетливый желобок, который, как полагают, является наружным слуховым проходом, этот желобок ведет сверху вниз к челюстному суставу. Затылок широкий, возможно, для прикрепления очень массивной туловищной мускулатуры; латерально он сливается с концами очень высоких скуловых дуг. Затылочная поверхность черепа является почти сплошной пластинкой, состоящей из соединенных затылочных и верхнеушных костей вместе с лежащими выше кожным верхнезатылочными и табличатыми костями. Затылочный мыщелок разделился и представляет собой двойное образование.
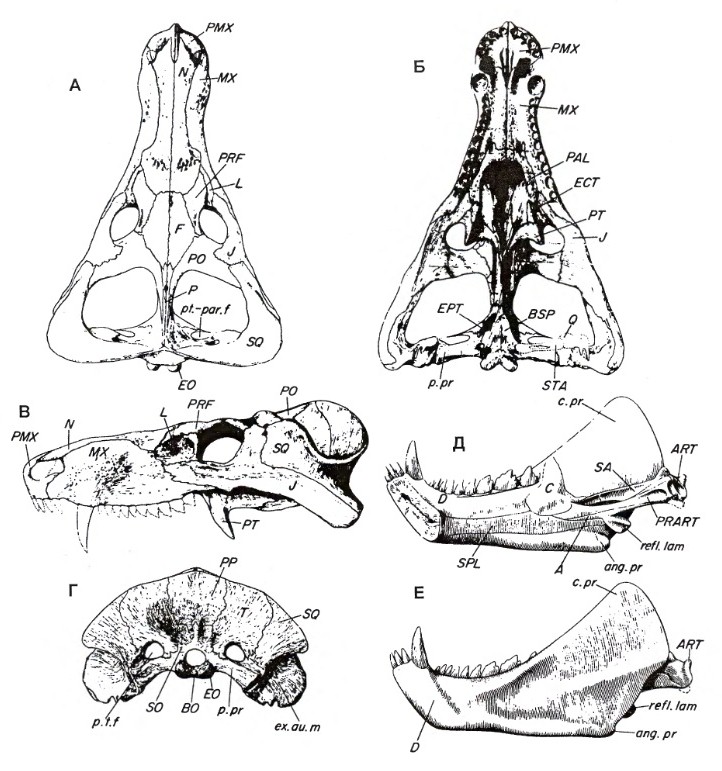
Прорисовки реконструированного черепа с нижней челюстью циногната (Cynognathus crateronotus): череп (А, Б, В, Г) и нижняя челюсть (Д, Е).
Продолжениями предчелюстных, верхнечелюстных и небных костей образована пластинка, которая лежит горизонтально ниже прежнего уровня крыши рта, так что образуется полное вторичное небо. Хоаны перемещаются назад через пространство, находящееся над вторичным небом. Оба предсошника срастаются в единое образование, от которого по срединной линии отходит отросток. Небные зубы отсутствуют. Межптеригоидных ям тоже уже нет, поскольку оба птеригоида срослись друг с другом в одно сплошное образование под передним концом черепной коробки; парасфеноид сохраняется в своем старом положении выше них и между ними. Птеригоиды укоротились и больше уже не протягиваются назад к квадратным костям, хотя расположенные выше них эпиптеригоиды могут продолжаться назад до этих последних. Сохраняются уменьшенные поперечные отростки птеригоидов. В дополнение к обычным костям черепной коробки эпиптеригоид образует плоскую пластинку, примыкающую к черепной коробке впереди переднеушной кости, а в более передней части имеется, по-видимому, пара костей, соответствующая орбитосфеноидам млекопитающих.
Этапы портретной реконструкции циногната (Cynognathus crateronotus). Автор — Р. Евсеев.
Зубная кость составляет значительно большую часть нижней челюсти, чем у более примитивных цинодонтов. Внешне она напоминает таковую крупных хищных млекопитающих. Назад и вверх от зубной кости отходит массивный венечный отросток с выпуклым задним краем, почти касающимся чешуйчатой кости. Яма жевательной мышцы глубокая, заходит далеко вперед. Нижний край зубной кости продолжается назад угловым отростком. Нижне-задний угол зубной кости выступает назад от края нижней челюсти. Задние кости нижней челюсти значительно редуцированы и объединены в узкий стержень, вставленный в желобок на медиальной поверхности зубной кости. Он состоит из сочленовной, предсочленовной, угловой и надугловой костей. Венечная кость остается в виде плоской пластины, прикрывающей передний конец этого стержня. Тип челюстного состава такой же, как у примитивных терапсид; он образован сочленовной и квадратной костями. Стремя тянется в виде тонкой палочки от отверстия внутреннего уха к квадратной кости.
Ямы и каналы на лицевой поверхности черепа указывают на концентрации нервов и кровеносных сосудов, которые могли быть связаны с вибриссами.
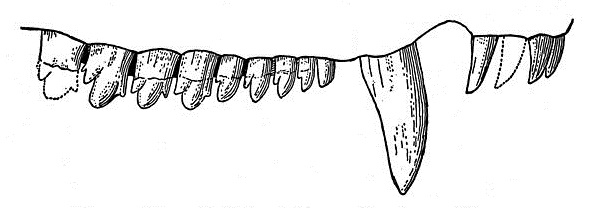
Прорисовка верхнего зубного ряда циногната (Cynognathus crateronotus), снаружи.
Зубы отчетливо дифференцированы, их формула: I4/i3, C1/c1, Pc9-10/pc9. Резцы и крупные клыки конические, несколько загнутые назад. Заклыковые зубы сжатые латерально, грубо пильчатые. Большинство задних заклыковых зубов трехзубчатые, с более мощными, чем у Thrinaxodon, зубцами и отклоненной назад главной вершиной, но на самых задних зубах появляются дополнительные вершины, расположенные позади главной. Лингвальный поясок на заклыковых зубах не развит. Обычно его отсутствие считают вторичным явлением, связанным с питанием циногната более крупной добычей. Предпоследний зуб увеличен и аналогичен хищническому зубу плотоядных млекопитающих. Наблюдается некоторое истирание зубов, но точного прикуса между верхними и нижними зубными рядами нет.
Щечной зуб циногната (Cynognathus crateronotus).
Посткраниальный скелет весьма схож с таковым у
Thrinaxodon, но отличается пропорциями, связанными с увеличением веса тела. Телосложение довольно массивное. Туловище вытянутое. В позвоночнике исчезли интерцентры, хотя тела позвонков остаются амфицельными. Насчитывается 6-7 шейных, 17-18 грудных, 4-5 поясничных и 3-4 крестцовых позвонка. Свободно подвижные двуглавые ребра имеются на всем протяжении от шеи до основания хвоста, постепенно укорачиваясь кзади. На задних спинных ребрах сохраняются расширенные пластинки, характерные для
Thrinaxodon. Сохраняются гастралии.
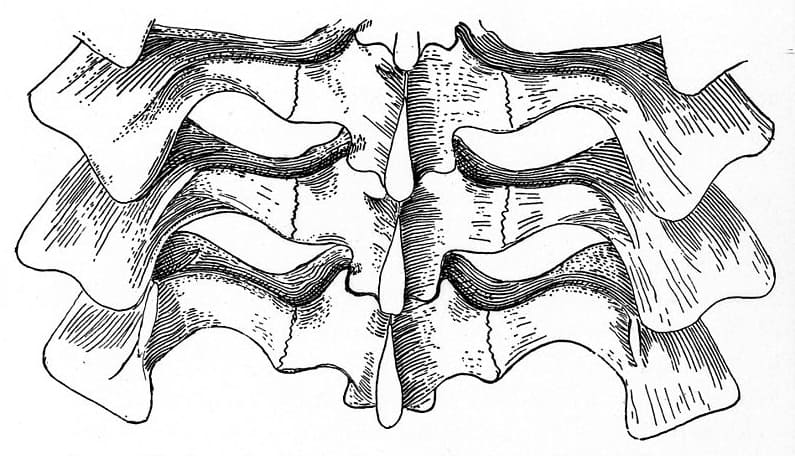
Прорисовка спинных позвонков циногната (Cynognathus crateronotus).
Плечевой пояс продолжает состоять из кожного пояса, в который входят межключица и ключицы (клейтрум исчез), и первичного пояса из лопатки и 2 коракоидных элементов. Передний край лопатки отвернут кнаружи (начало возникновения гребня, свойственного млекопитающим), и ключица прикрепляется к отростку, выступающему у нижнего конца гребня. В тазовом поясе подвздошная кость вытягивается скорее вперед, чем назад (последнее положение является примитивным), в связи с изменением положения мышц, и прикрепляется к возросшему числу крестцовых ребер. Лобковая и укороченная седалищная кости передвинулись назад (что также связано с изменением в положении мышц); между ними появилось хорошо развитое запирательное отверстие, которое частично разделяет их.
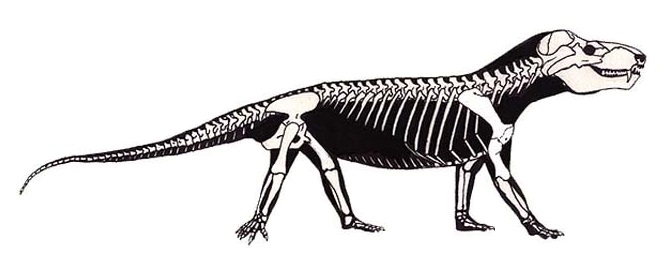
Прорисовка реконструированного скелета циногната (Cynognathus crateronotus), вписанного в телесные очертания.
Передние конечности несколько расставлены в стороны, а задние размещаюися непосредственно под телом. На бедренной кости головка развивается скорее сбоку, чем на конце этой кости, в связи с тем, что она повернута кпереди; а на наружной стороне имеется большой трохантер для мускулов, которые идут от подвздошной кости. Пальцы, за исключением I-го, имеют почти одинаковую длину в связи с тем, что конечностям свойственно передне-заднее движение. Фаланговая формула составляет 2 - 3 - 4 - 5 - 3, так как в III-м и в IV-м пальце сохраняются рудименты лишних фаланг.
Кортекс длинных костей конечностей выраженно зонален, построен из фиброламеллярной ткани и богато васкуляризирован, тогда как ламеллярная ткань в нем отсутствует. Это свидетельствует о непрерывности роста костной ткани.
Циногнаты (Cynognathus crateronotus) у туши крупного дицинодонта каннемейерии (Kannemeyeria simocephalus). Автор — М. Халлетт.
Циногнат, несомненно, являлся плотоядным. Он мог охотится на достаточно крупную добычу, но при случае не брезговал и падалью. Судя по всему, он являлся одним из наиболее грозных и подвижных хищников своего времени. Животное оказалось настолько успешным, что существовало на протяжении длительного времени и распространилось на обширных пространствах.
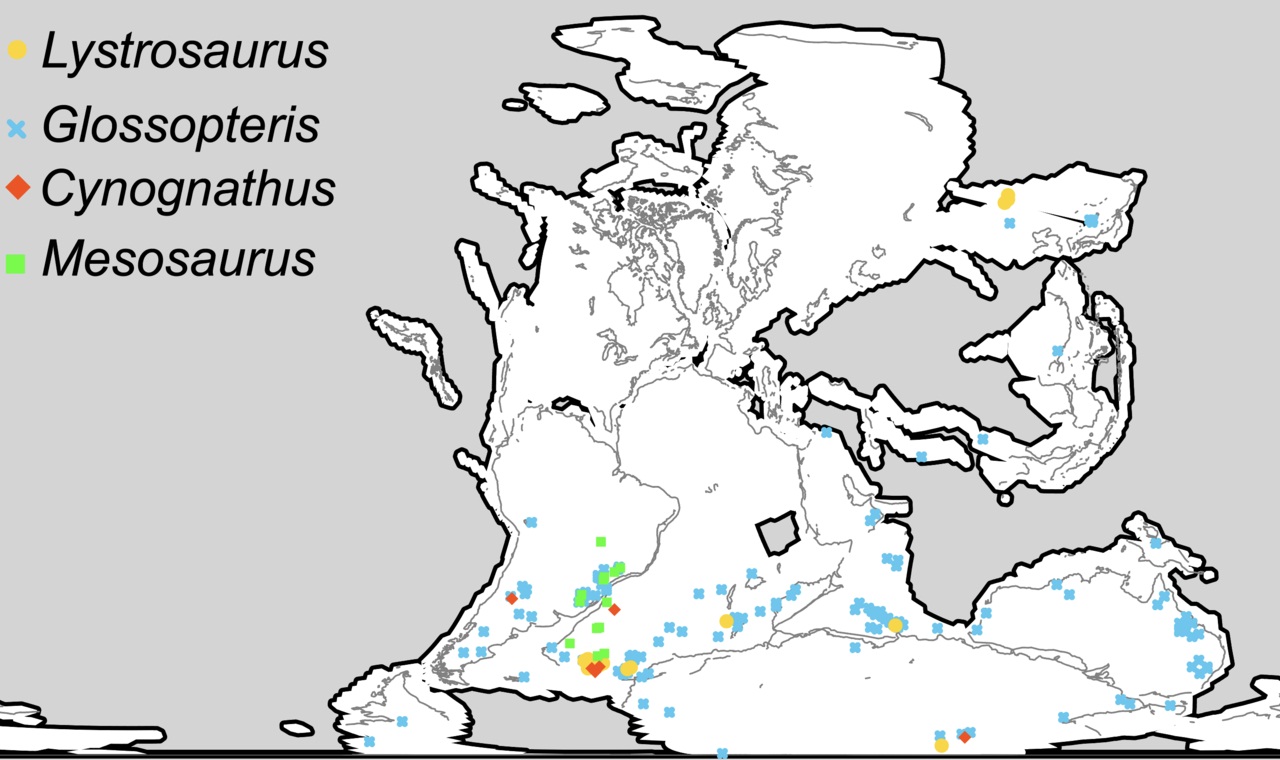
Карта некоторых находок ископаемых остатков
Cynognathus (отмечены красным цветом).
Ссылки
Новиковская А. Циногнат, наследник палеозоя. — 2023 (Элементы.Ру)
перейти Cynognathus (Paleobiology Database)
перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Иванов А. О., Черепанов Г. О. Ископаемые высшие позвоночные. — СПб.: Издательство Санкт-Петербургского университета, 2001
Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1 / Отв. ред. Ивахненко М. Ф. и Курочкин Е. Н. — М.: ГЕОС, 2008
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Очев В. Г. Еще не пришли динозавры. — Саратов: Научная книга, 2000
Павлова М. В. Палеозоология. Ч. 2. Позвоночные. — М.-Л.: Государственное издательство, 1929
Протеро Д. Отпечатки жизни. 25 шагов эволюции и вся история планеты. — СПб.: Питер, 2016
Ромер А. Ш. Палеонтология позвоночных. — М. — Л.: Государственное научно-техническое издательство нефтяной и горно-топливной литературы, 1939
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГеОС, 2009