Дицинодонты — Dicynodontia
Класс †Синапсиды (Synapsida)
Подкласс †Терапсиды (Therapsida)
Надотряд †Динообразные (Dinomorpha)
Отряд †Аномодонты (Anomodontia)
Подотряд †Хайнозавры (Chainosauria)
Инфраотряд †Дицинодонты (Dicynodontia)
Дицинодонты (Dicynodontia Owen, 1859) — инфраотряд подотряда хайнозавров (Chainosauria von Nopsca, 1923), происходящий из средней перми - ранней юры Африки, поздней перми - среднего или позднего триаса Мадагаскара, средней перми - позднего триаса Южной Америки, раннего - позднего триаса Антарктики, поздней перми - позднего триаса Европы, поздней перми - среднего триаса Азии, среднего - позднего триаса Северной Америки и, возможно, поздней перми - раннего триаса Австралии. Самая многочисленная группа синапсид: установлено порядка 14 семейств и 120 родов.
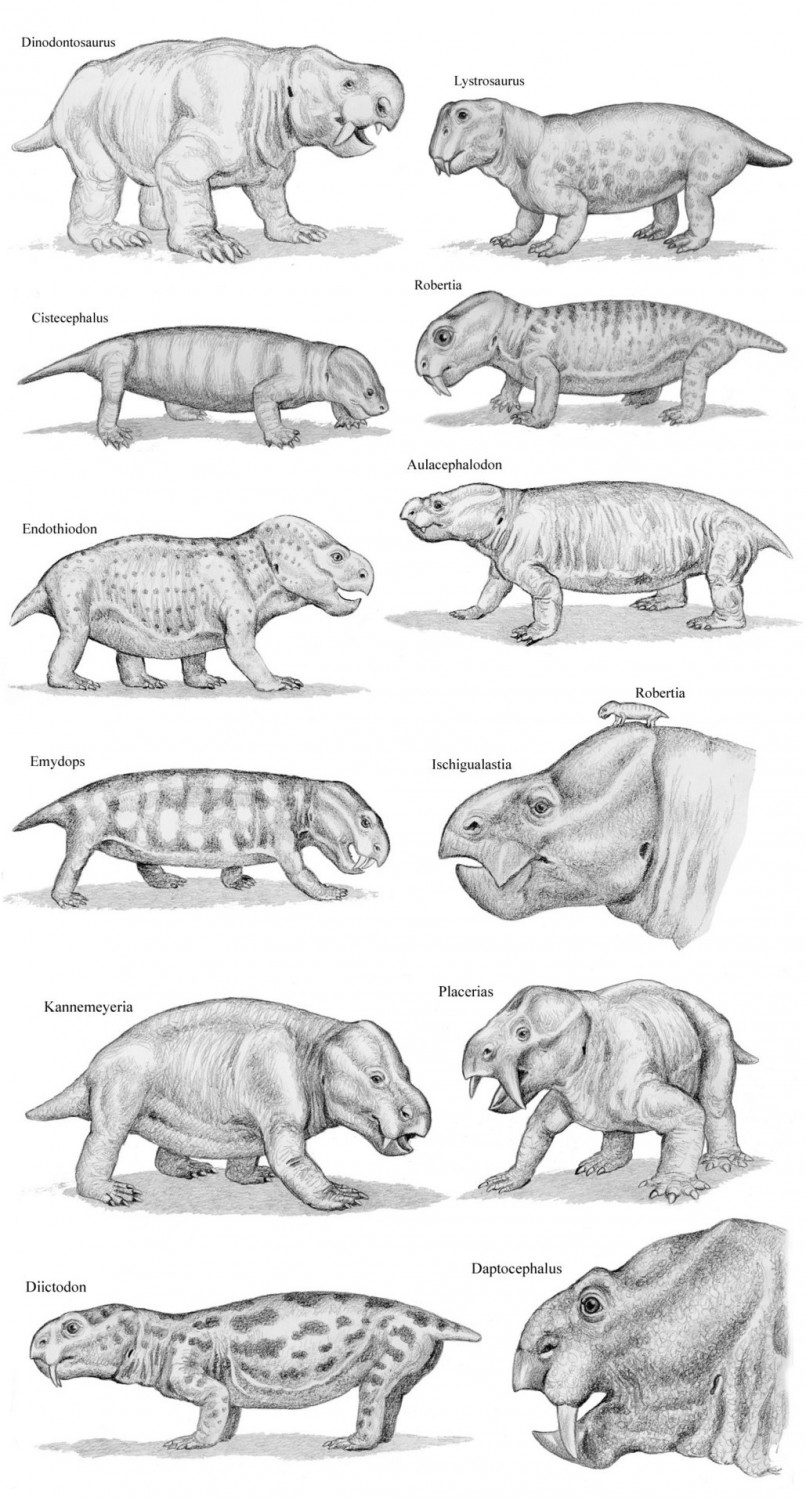
Реконструкции внешнего облика некоторых триасовых дицинодонтов (Dicynodontia), слева направо: Dinodontosaurus, Lystrosaurus, Angonisaurus, Ischigualastia, Myosaurus, Kombuisia. Автор — Г. Угуэто.
Название группы переводится как "двусобакозубые", или "двуклыкозубые", что указывает на специфическую особенность строения зубного аппарата ее типичных представителей. Это венец развития аномодонтной линии синапсид.
Размеры колеблятся от мелких до очень крупных: длина черепа варьирует приблизительно от 4 см до 1 м, общая длина — от 20 см до 4,5 м. Для дицинодонтов особенно характерно, что на протяжении длительной эволюционной истории они претерпели довольно незначительные морфологические изменения.
Смонтированный скелет листрозавра (Lystrosaurus murrayi).
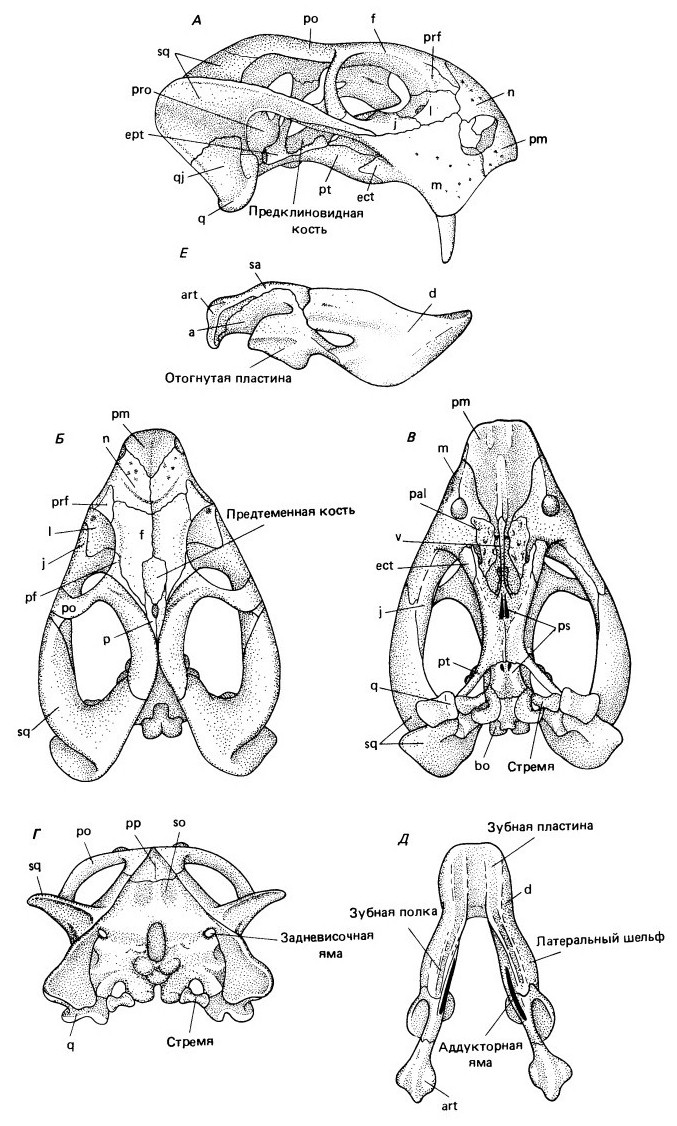
Череп обнаруживает значительную специализацию. Он массивный, укороченный в лицевой, удлиненный и расширенный в заглазничной области. У многих относительно мелких форм череп резко сужается в ростральной области. Ноздри расположены спереди над загнутыми межчелюстными костями. Глазницы направлены в стороны. Иногда имеется кольцо склеротики. Наверху височное окно, как и у горгонопид, ограничено длинным задним отростком заглазничной кости, смыкающейся у заднего края височного окна с чешуйчатой костью. Надвисочная кость утрачена. Височные окна очень большие и удлиненные.
Глубоко вогнутая снизу скуловая дуга широко выступает в сторону и назад, освобождая место для прохождения частей наружного нижнечелюстного аддуктора (m. adductor mandibulae extemus). Чешуйчатая и квадратно-скуловая кости, широко разрастаясь ниже заднего основания скуловой дуги, предоставляли место для отхождения дополнительного латерального пучка наружного аддуктора. Передний отросток чешуйчатой кости заходит под глазницу, а задний отросток скуловой кости редуцируется, из-за чего создается впечатление, что скуловая дуга переходит впереди в подглазничную.

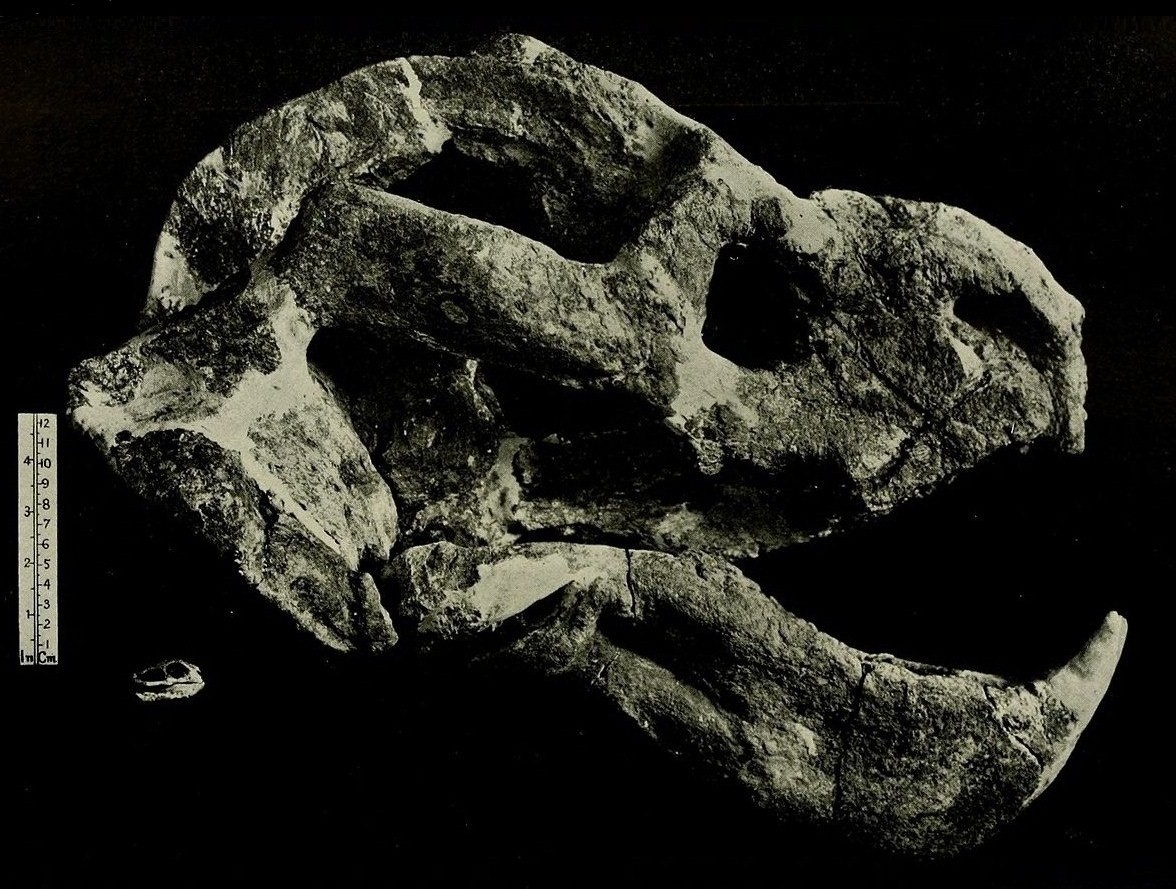
Череп с нижней челюстью Lystrosaurus.
Дицинодонты отличаются от венюковиоидов удлинением височной области и значительным развитием чешуйчатой кости, что приводит к образованию широкой костной пластины на латеральной поверхности черепа. Массивная чешуйчатая кость обладает широким отростком, нисходящим к области челюстного сочленения, и длинным передним отростком, входящим в скуловую дугу. Как и у венюковиоидов, на чешуйчатой кости имеется площадка для прикрепления наружного нижнечелюстного аддуктора.
Квадратная кость широкая, но короткая, срастается с редуцированной квадратно-скуловой и обычно рыхло соединяется с чешуйчатой костью. Межглазничный отдел черепа сравнительно высокий, теменной — узкий. Заднелобные кости могут исчезать. Теменные кости часто образуют сагиттальный гребень. Как у горгонопид, обычно имеется предтеменная кость, развитая преимущественно впереди пинеального отверстия, но обычно пинеальное отверстие полностью ограничено этой костью. Пинеальное отверстие в большинстве случаев крупное, иногда сильно уменьшенное (у Cistecephalidae). В некоторых случаях оно обращено при нормальном положении черепа не вверх, как, казалось бы, должно быть, а вперед.
Черепа с нижними челюстями дицинодонтов эндотиодона (
Endothiodon bathystoma) (большой) и эмидопса (
Emydops arctatus) (маленький).
Затылок широкий; поверхность его увеличивается за счет разросшихся чешуйчатых костей. На затылке имеются таблитчатые и непарная заднетеменная кости. Верхнезатылочная кость очень широкая. Задневисочное окно небольшое, полностью ограниченное верхнезатылочной и чешуйчатой костями. Порой сверху на черепе имеются костные шишки, которые могли служить основаниями рогов.
Кость в передней части верхней и нижней челюстей покрыта крошечными питающими отверстиями, напоминая поверхность, лежащую под роговым клювом птиц и черепах. Логично допустить, что у дицинодонтов имелся сходный покров, формирующий клювоподобную структуру с острыми краями.
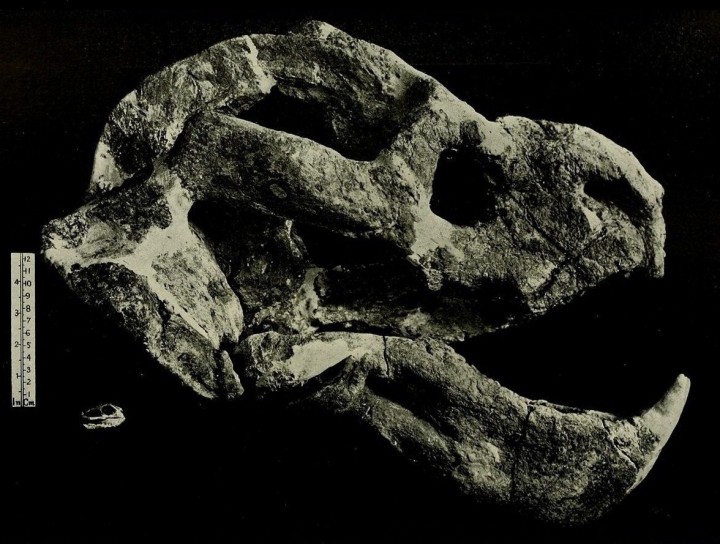
Прорисовки черепа и нижней челюсти Dicynodon: А — сбоку; Б — сверху; В — с неба; Г— сзади; Д — нижняя челюсть сверху.
Предчелюстные кости и сошники почти всегда срастаются по средней линии, лишь у самого примитивного семейства Eodicynodontoidae они остаются парными. Предчелюстные и верхнечелюстные кости, дополненные кнаружи от хоан отростками небных костей, формируют достаточно длинное вторичное небо. Отростки небных костей образуют "подушечки" с морщинистой поверхностью. Хоаны открываются позади средней точки черепа. В задней части вторичного неба образуется срединный костный гребень, переходящий в сошниковый гребень, разделяющий хоаны. В самой передней части небных отростков предчелюстных костей обычно имеются короткие парные гребни. Область крыловидной кости, служащая для отхождения крыловидной мышцы, разрастается вперед до верхнечелюстной кости, а не латерально, поэтому поперечные отростки крыловидных костей, бросающиеся в глаза у венюковиоидов, более не заметны. В задней части неба крыловидные кости срастаются с основной клиновидной костью. Небная область продвинута в связи с наличием шовного соединения между мозговой коробкой и крыловидными костями, заменившего подвижное базикраниальное сочленение.
Нижняя челюсть высокая и короткая. Ее симфиз массивный, желобчатый и очень длинный. У продвинутых форм имеются длинные вентральные валики на предчелюстных костях и плоские дентальные площадки на нижнечелюстном симфизе. Их наличие связано с развитием рогового чехла, покрывавшего неозубленные десна и переднюю часть неба. Как у венюковиоидов, на задней части зубной кости развит латеральный шельф. Зачастую сохраняется латеральное мандибулярное окно. Наружная пластина угловой кости, по крайней мере у некоторых дицинодонтов, округло отходит латерально, и полость под ее внутренней поверхностью необычно расширяется. В меньшей степени эта расширенность выражена и у венюковиоидов. Венечный отросток утрачен.
Распиленный верхнечелюстной клык динодонтозавра (Dinodontosaurus tener).
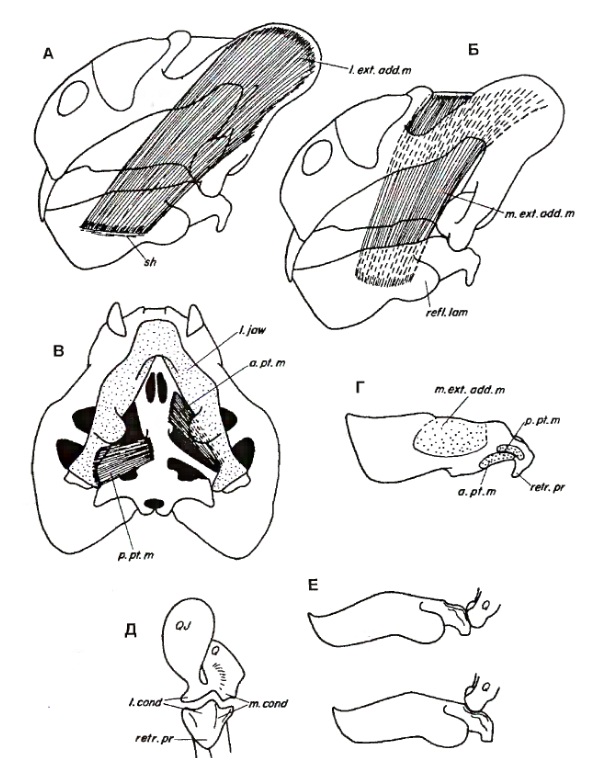
Форма челюстного сустава значительно отличается от свойственной венюковиоидам. Сочленовные мыщелки асимметричные, латеральные много шире медиальных. Суставная поверхность сочленовной кости почти вдвое длиннее, чем квадратной. У обоих мыщелков она выпуклая, если смотреть сбоку, но с продольной бороздой, что указывает на поступательное движение суставной поверхности нижней челюсти по краю квадратной кости в отличие от простого шарнирного сустава большинства тетрапод. Форма челюстей и височной области говорит о том, что основные открывающие и закрывающие рот мышцы были ориентированы почти горизонтально, т. е. большое значение имели передне-задние (пропалинальные) движения нижней челюсти. При закрывании рта нижняя челюсть "отъезжала" назад.
Зубы дицинодонтов в той или иной степени редуцированы. Передние концы челюстей лишены зубов всегда. В верхней челюсти обычно сохраняется только пара более или менее развитых бивнеобразных клыков, каждый из которых сидит на массивном клыковом выступе в передней части верхнечелюстной кости. Клыки круглые в сечении, цилиндрические, лишены эмали и обладают широко открытым корнем, обеспечивающим постоянный рост. Клыки слабо изогнуты кзади, в норме имеют лабиальные площадки стирания, видимо, о роговой чехол нижней челюсти. Обычно хорошо выражена и стертость вершин о субстрат, имеются даже экземпляры с прижизненно сломанными вершинами, заглаженными последующим употреблением. В ряде случаев верхние клыки свойственны лишь самцам.
Реконструкция черепной мускулатуры дицинодонта.
Иногда и верхние клыки исчезают, как у мелких, преимущественно позднепермских и раннетриасовых дицинодонтов, так и y очень крупных средне- и позднетриасовых. Как правило, вместо утраченных клыков развиваются клыкоподобные выросты верхнечелюстных костей. У примитивных форм на челюстях, помимо верхних клыков (которые, впрочем, иногда отсутствуют), сохраняются заклыковые зубы в виде однообразных мелких колышков, обычно смещенные внутрь от альвеолярного края кости. Такое смещение сильнее выражено в верхней челюсти, где щечные зубы расположены явно внутрь от клыка. Судя по всему, к редукции челюстных зубов привело совершенствование рогового чехла, функционально их заменившего.
У аномодонтов развивается новая крупная мышца. Уже у венюковиоидов наружный аддуктор нижней челюсти был на большом протяжении открыт снаружи. На латеральной поверхности их нижней челюсти имеется выемка, в которую заходила поверхностная часть этой мышцы. Место ее отхождения, возможно, находилось в процессе перемещения с медиальной поверхности задней части скуловой дуги на латеральную поверхность чешуйчатой кости. Сильно разросшаяся наружу чешуйчатая кость дицинодонтов обеспечила очень большую площадь отхождения увеличившейся мышцы. На латеральной поверхности нижней челюсти развилась специальная зона для ее прикрепления — боковой уступ.
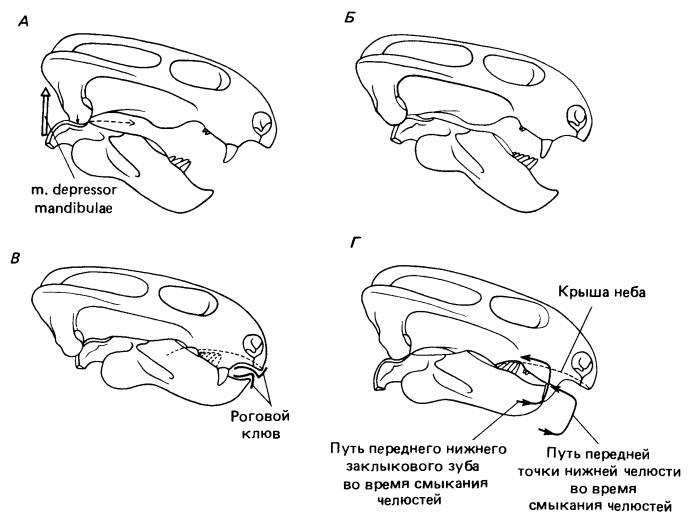
Схематичное изображение функций челюстного сустава дицинодонтов на примере Dicynodon: А, Б — вид сбоку черепа с заштрихованными поверхностным (А) и глубоким пучками (Б) поверхностного нижнечелюстного аддуктора; В — вид с неба глубоких пучков нижнечелюстного аддуктора; Г — нижняя челюсть снаружи с показом прикрепления аддукторных мышц; Д — челюстной сустав, вид сзади; Е — вид сбоку нижней челюсти в протрагированном (вверху) и ретрагированном (внизу) положениях относительно квадратной кости.
Наличие этой латеральной мышцы, по-видимому, уравновешивало более медиально направленную силу остальных элементов аддукторной мускулатуры. В результате стала возможной редукция поперечного выступа крыловидной кости, который у более примитивных синапсид предупреждает медиальное смещение нижней челюсти. Смена ориентации этого выступа увеличила площадь подвисочного окна и позволила распространяться вперед мускулам-аддукторам. Параллельно с этими изменениями уменьшается роль крыловидной мускулатуры.
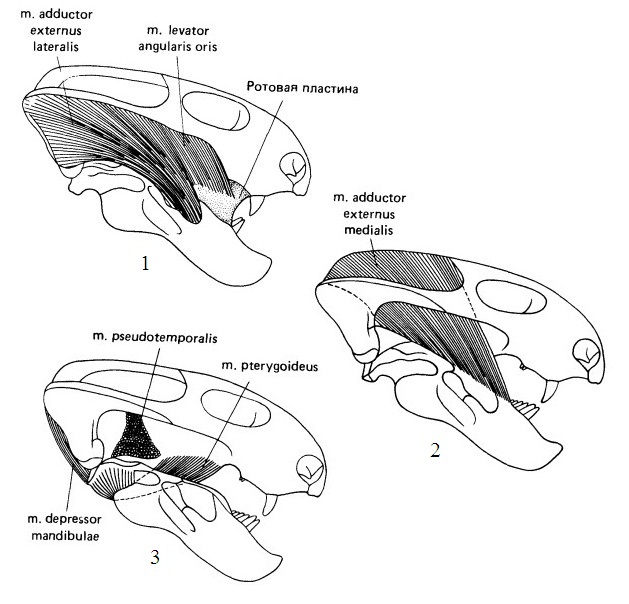
Реконструкция смыкающих челюсти мускулов на все более глубоких уровнях у примитивного дицинодонта эмидопса (Emydops arctatus).
Преобразования черепа и челюстной мускулатуры дицинодонтов во многом связаны с изменениями их механизма питания. Корм обрабатывался посредством передне-задних нарезающих движений нижней челюсти. Установлено, что наибольшая раздавливающая пищу сила возникала при ретракции нижней челюсти, когда рот дицинодонтов был почти закрыт. Открывался он из ее максимально оттянутого назад положения. В начале смыкания челюстей нижняя занимает переднее положение, подтягиваясь вперед крыловидной мышцей. Затем смыкание и скользящее оттягивание ее назад шли одновременно под действием аддукторов, отходящих от задней части височной области. Прижимание нижней челюсти к небу обеспечивалось вращением артикулярного сочленовного мыщелка относительно квадратного.
У примитивных дицинодонтов такое движение приводит к разрезанию пищи в 2 контактных зонах: сзади — между оставшимися щечными зубами нижней челюсти, верхнечелюстной костью и небом, и спереди — между симфизной пластинкой нижней челюсти челюсти и клыковым выростом верхней. Эти поверхности были покрыты роговым веществом, по-видимому, с острыми режущими кромками. У продвинутых форм, которые полностью утратили щечные зубы, разрезание ограничивалось передней областью, площадь поверхности которой увеличилось. Ни перетирания, ни пережевывания пищевых объектов не происходило; измельчение шло исключительно за счет режущих движений острых краев клюва и продольных гребней на вторичном небе, несущих роговое покрытие.
Стадии жевательного цикла примитивного дицинодонта эмидопса (Emydops arctatus): А — опускание и начало выдвигания вперед (пунктирная стрелка указывает направление движения нижней челюсти); Б — полное выдвигание и начало поднятия; В — укус клювом и начало ретракции; Г — полная ретракция.
В то время как череп дицинодонтов весьма постоянен в своих пропорциях, что обусловлено уникальной специализацией ротового аппарата, их посткраниальный скелет в значительной степени различается в зависимости от веса тела и образа жизни. Шея короткая. Туловище в большей или меньшей степени удлиненное, но широкое, бочкообразное. Конечности короткие и массивные. Постановка конечностей у мелких и средних по размеру дицинодонтов широко расставленная. У крупных триасовых форм расставленные в стороны передние конечности остро изогнуты в локте, а постановка задних конечностей приближается к парасагиттальной плоскости. Хвост короткий, резко отграниченный от туловища.
Обычно имеется 26-28 (до 30) предкрестцовых позвонков, из них порядка 7 шейных. Поясничный отдел позвоночника не выражен. В крестце у мелких и средних по размеру форм 3-4 позвонка, у крупных триасовых родов их число возрастает до 5-8 за счет присоединения задних спинных позвонков. Тела позвонков глубокоамфицельные, укороченные; интерцентры отсутствуют.
Следовые отпечатки дицинодонта из триаса провинции Мендоса, Аргентина.
Плечевой пояс с широкой лопаткой и с акромиальным отростком. Иногда сохраняется клейтрум. Грудина развита и обычно хорошо окостеневает. Подвздошная кость наклонена вперед, лонная и седалищная — назад. Подвздошная кость расширенная, но тазовый пояс, по-видимому, без симфиза. Лонная кость зачастую относительно маленькая. Плечевая кость массивная, без эктэпикондилярного отверстия. Ступни обычно широкие, с короткими пальцами и мощными когтями. Фаланговая формула редуцирована до маммальной: 2 - 3 - 3 - 3 - 3. У высокоспециализированных роющих Cistecephalidae кисть сужена за счет редукции крайних пальцев.
Вероятно, тело было покрыто мягкой кожей, может быть, иногда с зачатками шерсти. Мелкие ямки на морде порой трактуются в качестве отпечатков оснований вибрисс. Дицинодонты размножались, откладывая яйца в норах или кучах гниющей растительности. Высокий уровень содержания в трубчатых костях фиброламеллярной костной ткани указывает на быстрый рост в ювенильном возрасте с последующим его замедлением.
Предполагаемые яйца дицинодонтов из поздней перми Южной Африки.
Несмотря на свойственную дицинодонтам морфологическую консервативность, обусловленную универсальностью приобретенных эволюционных адаптаций, они сильно различались экологически. Сегодня все представители группы обычно рассматриваются как в достаточной мере наземные животные. Сочетание заместившего зубы клюва, сильной челюстной мускулатуры и челюстного мыщелка "открытого" типа, обеспечивавшего свободные возвратно-поступательные движения челюстей, позволило создать достаточно универсальный челюстной аппарат, схожий с таковым современных черепах и допускавший столь же широкий спектр потребляемой пищи.
О пищевых предпочтениях дицинодонтов споры ведутся до сих пор. Разные авторы предполагали для них питание плодами, грибами, членистоногими. Существовала даже гипотеза о преимущественной падалеядности дицинодонтов. Разумеется, она не исключена для многих фитофагов (например, современных свиней), однако облигатное падалеедение все же представляется крайне маловероятным. Судя по всему, некоторые мелкие формы действительно питались беспозвоночными или были всеядными, но большинство дицинодонтов представляли собой в большей или меньшей степени специализированных растительноядных. Судя по широкому разбросу в общих размерах и специфических отличия черепного строения, рацион у представителей разных линий должен был иметь свои особенности.
Реконструкция дицинодонтов Dicynodon, объедающих труп парейазавра Scutosaurus, выполненная А.П. Быстровым. Академик П.П. Сушкин, изучив найденного В.П. Амалицким на реке Северной Двине сравнительно небольших или средних размеров дицинодонтов, пришел к выводу, что крупные и подобные щипцам-кусачкам челюсти, могучая шея и крепкие передние ноги выдают в этих животных падалеядов. И в России, и в Южной Африке их находят вместе с крупными растительноядными ящерами парейазаврами и похожих на саблезубых тигров горгонопидами. Сушкин полагал, что горгонопиды охотились на парейазавров, а дицинодонты, таким образом, могли являться третьим членом этого сообщества и подъедали за хищниками остатки их добычи.
В подтверждение того, что фитофагия являлась генеральным эволюционным трэндом дицинодонтов, прежде всего свидетельствует обилие их остатков во многих местонахождениях, что прямо отображает многочисленность этих животных в биоценозах. Подобная обильность по сравнению с явными хищниками говорит о низком положения дицинодонтов в трофических пирамидах животных сообществ и соответствует многочисленности растительноядных в современных экосистемах. Во многих сообществах поздней перми и раннего триаса эти синапсиды были доминирующими фитофагами. Так, в позднепермских отложениях бассейна Карру на юге Африки из каждых найденных 10 черепов обычно чуть ли не 9 принадлежат дицинодонтам. С другой стороны, в Северодвинском местонахождении остатки дицинодонтов по сравнению с парейазаврами немногочисленны.
Не исключено, что дицинодонты стали первыми крупными фитофагами, которые не только отыскивали и перерабатывали пищу, но и активно добывали ее, возможно, выкапывая из почвы. Это позволило питаться относительно мягкими корнями имевшихся повсеместно растений, тех же членистостебельных, надземные части которых чрезвычайно жестки, а молодая поросль и генеративные органы по количеству не могли удовлетворять крупных растительноядных. Усовершенствовав со временем челюстной аппарат, дицинодонты смогли включить в рацион и ксерофильные жестковолокнистые растения с высоким уровнем содержания целлюлозы. Это обеспечило широкое распространение и длительное процветание группы. Ее представители были самыми многочисленными, разнообразными и дольше прочих существовавшими аномодонтами.
Реконструкция возможного способа питания триасовых дицинодонтов каннемейероидов, выполненная Артуром Ричардом Ивором Круикшенком. По мнению этого исследователя, каннемейероиды могли выворачивать лбом крупные растения или нагибать их когтистыми лапами. О такой способности этих животных, по мнению автора, свидетельствуют их сильная крыша черепа и крюкоподобные конечные фаланги пальцев. Круикшенк полагал, что каннемейероидов следует сравнивать с вымершими наземными ленивцами Нового Света. Строение нижней челюсти некоторых среднетриасовых дицинодонтов позволяет, по его мнению, предположить, что, как и ленивцы, они имели длинный язык для захвата ветвей и листьев.
Скользящий механизм движения нижней челюсти в сочетании с острым роговым клювом обеспечивал эффективное срезание и измельчение растительного материала дицинодонтами. Спорным остается вопрос о функциональности верхних клыков. Последние работы противоречат возможности их использования для рытья почвы и добывания корней и клубней. Исследование рабочих поверхностей клыков дицинодонтов из поздней перми Южной Африки показало, что лишь у некоторых их родов (Dicynodon, Lystrosaurus, Diictodon и, возможно, Dicynodontoides) и только при максимальных размерах клыков, даже при закрытой и полностью отведенной назад нижней челюсти, за пределы челюстей выступает не более 1/3 длины клыка. Наблюдаемая степень износа клыков также не подтверждает их активного использования для копания грунта.
Одним из возможных объяснений функционального значения дицинодонтовых клыков-бивней может являться предположение об использовании их в качестве специфического орудия для захвата и удержания кормового объекта в клещевом замке, образующемся при смыкании челюстей из верхних клыков (или замещающих их верхнечелюстных выростов) и нижней челюсти. Такой механизм мог оказаться весьма эффективным при питании, например, слабо и неглубоко укорененными побегами и корневищами, позволяя извлекать из почвы их крупные части для дальнейшей обработки в ротовой полости. Подобное копание могло осуществляется не клыками, а покрытой роговым чехлом передней частью морды в мягком и скорее всего влажном грунте. Данную роющую деятельность можно весьма приблизительно сравнить с таковой свиней. Она выглядит вполне уместной в околоводных и периодически затапливаемых ландшафтах с преобладанием плаунов, хвощей и папоротников. Помимо морды, копание в ряде случаев наверняка осуществлялась сильными конечностями, вооруженными большими когтями. Копательные адаптации конечностей, помимо Cistecephalidae, отчетливо выражены, например, у Diictodon.
Строение косточек склеротикального кольца указывает на дневной образ жизни подавляющего большинства дицинодонтов, хотя многие из них и жили в норах.
Естественная отливка норы дицинодонта, происходящая из отложений формации Наобаогоу в автономном районе Внутренняя Монголия, Китай.
По своему происхождению дицинодонты близки к дромазаврам, которые образуют по отношению к ним сестринскую группу. Дромазавры напоминают дицинодонтов общей конфигурацией черепа с широкой теменной (межвисочной) областью, но у них сохраняется задний отросток скуловой кости, менее развит передний отросток чешуйчатой, нет клыков и рогового клюва, зато сохраняются передние челюстные зубы. Поперечные отростки крыловидных костей у дромазавров редуцируются, небные зубы, как и у дицинодонтов, утрачены, но, самое важное, что у дромазавров мало развито вторичное небо и начинает выражаться комплекс преобразований, связанных с передне-задними "нарезающими" движениями нижней челюсти. Короноид у дромазавров обычно утрачен, как и у дицинодонтов. По крайней мере в известных случаях, седалищная кость таза дромазавров направлена назад от уровня подвздошной, а фаланговая формула, как и у дицинодонтов, маммального типа.
Д. Кинг (1988 г.) и М.Ф. Ивахненко (2003 г.) включают дромазавров и венюковиоидов в один инфраотряд с дицинодонтами, не выделяя последних как единую группу. Л.П. Татаринов (2009 г.) считает вполне обоснованным родство дромазавров с дицинодонтами, но предпочитает выделять последних в особую группу.
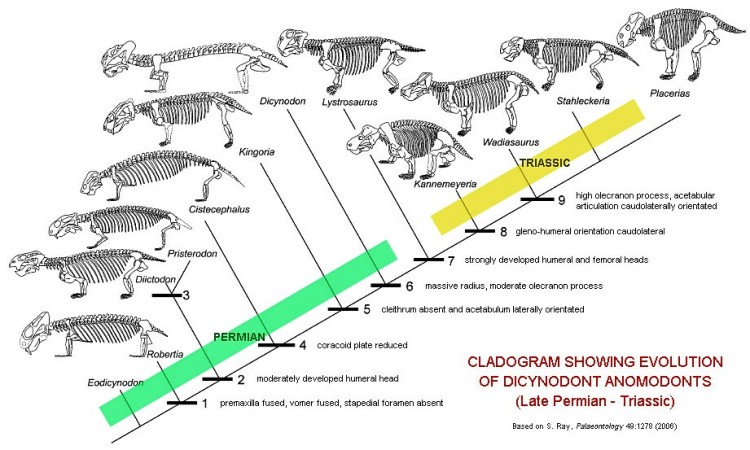
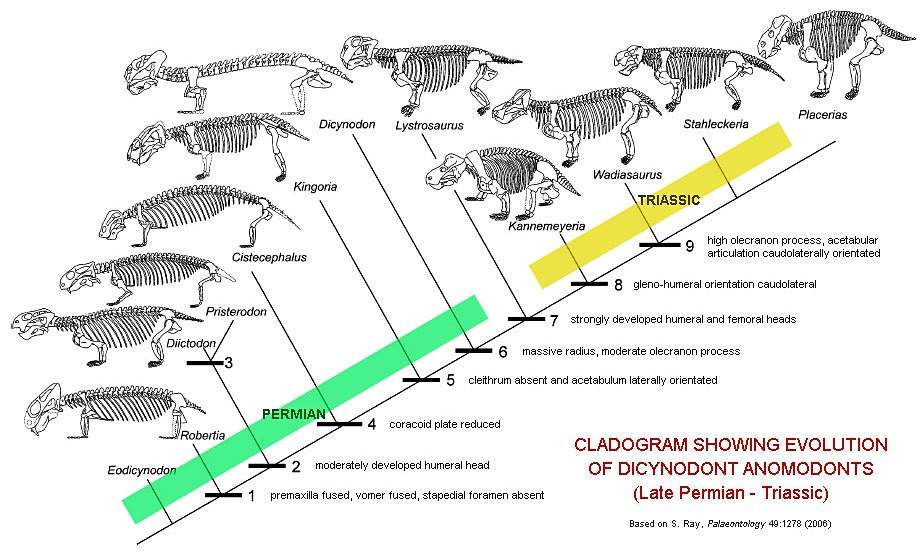
Основные вехи эволюции дицинодонтов (Dicynodontia).
Древнейшие известные дицинодонты происходят из среднепермских отложений Южной Африки (Eodicynodon, давший название одноименный биостратиграфической зоне). Таким образом, они появляются в геологической летописи раньше дромазавров. Основное направление в начальной эволюции группы — редукция челюстных зубов и замещение их специфическим клювом из сильно ороговевшей ткани. У многих пермских дицинодонтов еще сохранялись немногочисленные, тесно расположенные щечные зубы, как правило, не более 5 в верхней и не более 9 в нижней челюсти, но у рода Endothiodon число щечных зубов достигало нескольких десятков. Для большинства пермских родов характерно длинное туловище и короткие конечности с крупными когтями.
После вымирания диноцефалов в конце средней перми у дицинодонтов произошла мощная эволюционная вспышка, приведшая к их широкому распространению и взрывному росту разнообразия. В поздней перми дицинодонты были самыми многочисленными наземными позвоночными и доминирующими растительноядными в большинстве наземных экосистем. Особенно обильными они были Южной и Восточной Африки, откуда происходит до 60 родов; известны дицинодонты и в поздней перми Европы, Китая, Индии и США. Средние по размерам формы (Aulacephalodon, Endothiodon, Rhachiocephalus), возможно, вели полуводный образ жизни. Относительно мелкие дицинодонты жили в норах (Robertia, Diictodon) или вообще стали роющими (Cistecephalus, Kawingasaurus), о чем говорит клиновидная форма черепа, а также строение конечностей и их поясов. Но, в любом случае, пермские дицинодонты существенно уступали по размерам современным им парейазаврам.
Разнообразие дицинодонтов (Dicynodontia). Автор — В. ван дер Мерве.
Во время глобального биотического кризиса, разразившегося на исходе перми, большинство дицинодонтов вымерло. Уцелели лишь 2-3 рода, среди них — Lystrosaurus, животное средних размеров, распространившееся почти по всей Пангее. В триасовое время дицинодонты снова становятся многочисленными и распространяются по всем материкам, за исключением Австралии. Их относительно мало в среднем триасе Южной и Восточной Африки, но они обычны в Оренбуржье (Россия) и сохраняются также в Китае, Индии, Южной Америке и Антарктиде. Среди триасовых дицинодонтов преобладали крупные формы (Kannemeyeria, Stahleckeria) с пропорциями тела тяжеловесных растительноядных млекопитающих — например, носорогов. Из зубов у них остались лишь верхнечелюстные клыки, которые порой замещались клыкоподобными костными выступами.
В позднем триасе дицинодонты стали постепенно угасать, возможно, из-за конкуренции с ринхозаврами и растительноядными цинодонтами, а также бурной эволюцией охотившихся на них крупных архозавров. В эту эпоху дицинодонты были обычны преимущественно в Южной Америке; несколько родов сохранялось в США и в Марокко. Одними из последних представители группы были Lisowicia, Placerias и Ischigualastia (соответственно, поздний триас Европы, Северной и Южной Америк), достигавшие величины носорога. Последние достоверно установленные дицинодонты известны из ранней юры Африки. Фрагментарные остатки из Австралии, которые порой приписывали раннемыловым дицинодонтам, на поверку оказались остатками плейстоценового сумчатого Diprotodon.
Листрозавры (Lystrosaurus murrayi) в естественном окружении. Автор — З. Буриан.
В последние десятилетия ряд авторов предприняли ревизию системы дицинодонтов, используя и кладистические методы (Клювер и Кинг (1983 г.), Хопсон (1991 г.), Хопсон и Бархьюзен (1986 г.), Кинг (1988, 1990 гг.), Кинг и Рубидж (1993 г.), Кокс (1998 г.)), что способствовало уменьшению разногласий в их воззрениях на систему группы, но в целом система дицинодонтов еще не установилась. Т. Кемп (1982 г.), например, не выделяя надсемейственных групп, делит дицинодонтов на 7 семейств: Eodicynodontidae, Endothiodontidae, Kingoriidae, Diictodontidae, Dicynodontidae, Lystrosauridae и Kannemeyeriidae. Д. Кинг (1988 г.) выделяет в составе собственно дицинодонтов 5 надсемейств: Eodicynodontoidea, Endothiodontoidea, Pristerodontoidea (включая собственно дицинодонтид, листрозаврид и каннемейериид), Diictodontoidea (включая цистецефалид) и Kingorioidea. Б. Кокс (1998 г.) разделяет дицинодонтов на 5 надсемейств: Eodicynodontoidea, Robertoidea, Dicynodontoidea, Endothiodontoidea и Emydopoidea. Следует указать, что Д. Кинг не удалось поместить в выделенные таксоны семейственного ранга 20 родов.
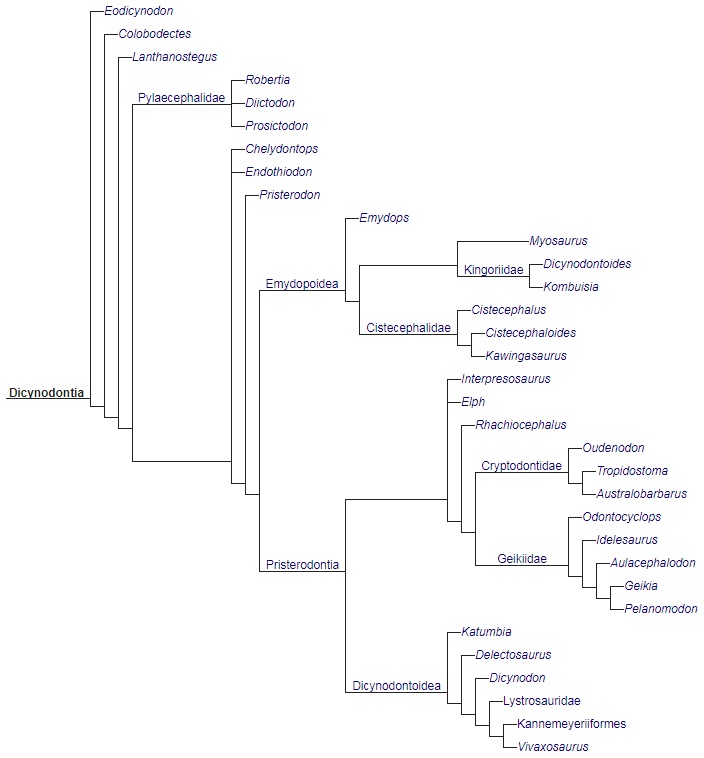
Кладограмма, иллюстрирующая родственные связи дицинодонтов (по Ангельчику и Рубиджу, 2010 г.).
Ссылки
Dicynodontia (Paleobiology Database)
перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Иванов А. О., Черепанов Г. О. Ископаемые высшие позвоночные. — СПб.: Издательство Санкт-Петербургского университета, 2001
Ивахненко М. Ф. Тетраподы Восточно-Европейского плаката — позднепалеозойского территориально-природного комплекса. — Пермь, 2001
Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1 / Отв. ред. Ивахненко М. Ф. и Курочкин Е. Н. — М.: ГЕОС, 2008
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Орлов Ю. А. В мире древних животных. 3-е изд. — М.: Наука, 1989
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Очев В. Г. Еще не пришли динозавры. — Саратов: Научная книга, 2000
Павлова М. В. Палеозоология. Ч. 2. Позвоночные. — М.-Л.: Государственное издательство, 1929
Протеро Д. Отпечатки жизни. 25 шагов эволюции и вся история планеты. — СПб.: Питер, 2016
Ромер А. Ш. Палеонтология позвоночных. — М. — Л.: Государственное научно-техническое издательство нефтяной и горно-топливной литературы, 1939
Сурков М. В. О трофических адаптациях жевательного аппарата позднепермских дицинодонтов. // Геология и минеральные ресурсы Европейского Северо-Востока России. Материалы XIV Геологического съезда Республики Коми. 13-16 апреля 2004 г. Т. 3. — Сыктывкар: Геопринт, 2004
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГеОС, 2009