Диноцефалы — Dinocephalia
Класс †Синапсиды (Synapsida)
Подкласс †Терапсиды (Therapsida)
Надотряд †Динообразные (Dinomorpha)
Отряд †Диноцефалы (Dinocephalia)
Диноцефалы (Dinocephalia Seeley, 1894) — отряд надотряда динообразных (Dinomorpha Ivakhnenko, 2003), происходящий из средней перми Европы, Азии, Африки и Южной Америки. Установлены 2 подотряда.
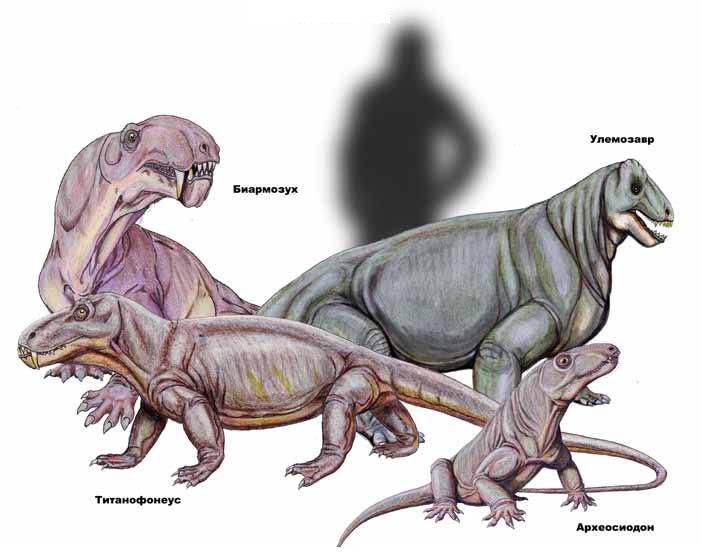
Разнообразие диноцефалов. Автор — Д. Богданов.
Синоним: Deinocephalia.
Свое название, которое переводится как «странноголовые», группа получила из-за свойственного многим ее представителям утолщения костей крыши черепа.
Разнообразные примитивные терапсиды, включающие как сравнительно мелких и подвижных, так и очень крупных тяжело сложенных животных. Общая длина варьирует от 1 до 5 м, вес — от нескольких килограммов до 2 т.
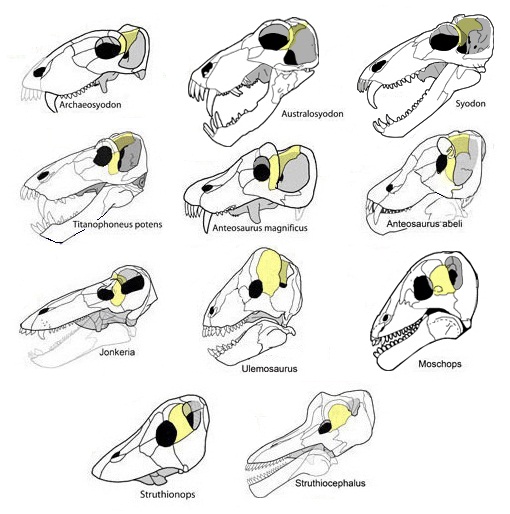
Прорисовки реконструированных черепов диноцефалов. Автор — Д. Петерс.
Череп высокий и очень массивный, длинный у хищных форм и относительно короткий у растительноядных. Морда обычно направлена чуть вниз. Ноздри с ее конца сдвинуты назад и разделены сильно удлиненными восходящими отростками премаксилл, вклинивающимися между носовыми костями. Носовые кости образуют длинные, направленные вверх отростки, глубоко вклинивающиеся между лобными. Глазницы обычно небольшие. Подглазничных отверстий нет. Предглазничные кости иногда, как и у пеликозавров, не соединяются с нисходящими отростками теменных.
Заглазничная часть черепа укороченная. Височное окно сравнительно небольшое, антеросинапсидного типа, развитое преимущественно в передне-верхнем направлении. При этом образуется антеродорсальное мускульное вдавление, ограниченное спереди гребнем заднего края заглазничной кости, смещенным в заглазничную область. Челюстная мускулатура в височном окне располагается практически вертикально. При продвижении ее передних пучков вперед, в надглазничную область крыши черепа, образуется мускульный блок. Заглазничная и чешуйчатая кости смыкаются по верхнему краю височного окна. Скуловая дуга несколько опущена. Верхняя часть затылочной пластины чешуйчатой кости вертикальная. Заднелобная кость всегда развита. Предтеменной кости нет. Теменные кости очень широкие.
Кости крыши черепа часто сильно утолщены (пахиостоз). Назначение утолщенных черепных костей остается неясным. Согласно самому распространенному предположению, животные использовали их во внутривидовых поединках (толкались лбами подобно баранам или морским игуанам). Альтернативная гипотеза — пронизанные кровеносными сосудами кости служили для терморегуляции. Крупное пинеальное отверстие расположено на высоком костном бугре и окружено валиком. Иногда имеется развитый сагиттальный гребень. Затылок всегда очень широкий, вогнутый, иногда резко наклоненный верхней частью назад. Затылочный мыщелок одинарный, часто выступающий вниз.

Черепной фрагмент южноафриканского диноцефала с чрезвычайно развитым каналом пинеального органа.
Вторичного неба нет. Хоаны расположены у переднего края небной поверхности, их отверстия сильно удлинены. Сошник парный, удлиненный, желобчатый. Птеригоиды сращенные, их квадратные ветви всегда хорошо развиты. Верхняя крыловидная кость узкая. Наружные крыловидные кости большие. На птеригоидах часто развиты зубы. Высокий носоглоточный желоб и очень мощные птеригоидные отростки на небе отсекали закрытую пасть от дыхательных путей. Внутреннее ухо соединяется с овальным окном длинным костным перилимфатическим каналом. Слуховая косточка (стремя) сохраняет массивность. Квадратноскуловая кость обычно широко выходит на наружную поверхность черепа. Область челюстного сочленения обычно опущена ниже уровня зубного ряда. Зубная кость относительно небольшая. Нижняя челюсть без венечного отростка.
У хищных диноцефалов зубная система дифференцирована, растительноядные формы четкой дифференциации не имеют. Нижнечелюстные резцы обычно заходят между верхнечелюстными. Как правило, резцы очень мощные и часто с пятками по внутреннему краю, увеличивающими их эффективность при пережевывании пищи. Такое необычное использование передних зубов иногда связывают с отсутствием вторичного неба, исключающим пережевывание пищи с помощью задних зубов без нарушения дыхания. Клыки иногда значительно развиты. Заклыковые зубы слабые.
Посткраниальный скелет массивный. Позвонки глубоко амфицельные, прободенные хордой. Пояса конечностей весьма примитивные, пластинчатые. В плечевом поясе обычно присутствует клейтрум. Лопатка без акромиона. Передние конечности несколько длиннее задних. Передние конечности широко расставлены, задние расставлены или более-менее прямые. Наклоненный назад затылок диноцефалов также свидетельствует о ходьбе на полувыпрямленных конечностях. Плечевая кость с обоими эпикондилярными отверстиями. Внутренний трохантер бедренной кости не развит. Кисти и стопы широкие, симметричные. Примечательно, что практически все известные в этом отношении диноцефалы имеют на маммальную фаланговую формулу: 2 - 3 - 3 - 3 - 3. По этому признаку они ближе к млекопитающим, чем горгонопиды и даже примитивные цинодонты.
Вероятно, диноцефалы имеют южногондванское происхождение. Они составляли основу среднепермских фаун наземных позвоночных на территории Европейской части России, Китая и Южной Африки. Диноцефалы населяли прибрежные леса, многие из них вели полуводный образ жизни. По характеру питания среди них выделяются хищные, всеядные и вторично растительноядные формы. Передвигаться диноцефалы могли только медленно — они не спеша ходили. Причины массивности, так ярко выраженной у многих представителей группы, окончательно не ясны; несомненно только то, что данная особенность имела какое-то приспособительное значение, вследствие чего и была закреплена естественным отбором. Эти животные вымерли в конце средней перми вследствие иссушения климата. Их сменили другие терапсиды.
Систематика
Первоначально остатки диноцефалов были описаны из Южной Африки, затем их распознали среди находок из Приуралья, а относительно недавно обнаружили в Китае и Бразилии. При этом следует отметить, что первые научно описанные фрагменты костей диноцефалов были получены из медистых песчаников Приуралья. С.С. Куторга, описавший их в 1838 г., полагал, что остатки принадлежат неполнозубым млекопитающим. В этом отношении он оказался ближе к истине, чем Р. Оуэн, который первоначально всех южноафриканских терапсид сближал с крокодилами.
Сечас практически общепринято разделение диноцефалов на более примитивных, преимущественно хищных, титанозухов (Titanosuchia) и специализированных растительноядных тапиноцефалов (Tapinocephalia). Зачастую титанозухов относят к предкам териодонтов, а тапиноцефалов — к предкам дицинодонтов, и иногда диноцефалы не выделяются как самостоятельная группа.
П.К. Чудинов (1983 г.) разделял титанозухов на 5 семейств: Brithopodidae, Deuterosauridae, Estemmenosuchidae, Anteosauridae и Titanosuchidae, а тапиноцефалов на 2 семейства: Tapinocephalidae и Styracocephalidae. М.Ф. Ивахненко (1995, 2003 гг.) сохраняет разделение диноцефалов на титанозухов и тапиноцефалов, но первоначально выделил в каждой из групп по 2 надсемейства: Titanosuchoidea и Deuterosauroidea в титанозухах и Tapinocephaloidea и Struthiocephaloidea в тапиноцефалах. К титанозухоидам М.Ф. Ивахненко относил и монотипическое семейство Biseridensidae, описанное из Китая Ж. Ли и З. Ченгом (1997 г.) в качестве эотитанозухид. Еще 2 китайских рода диноцефалов были отнесены описавшими их З. Ченгом и Ж. Ли (1997 г.) к примитивным антеозавридам и выделены в особое семейство Stenocybusidae.
Большие изменения в систему диноцефалов внесла Д. Кинг (1988 г.), приняв их в ранге подотряда аномодонтов и разделив на 2 надсемейства — Estemmenosuchoidea и Anteosauroidea. Estemmenosuchoidea рассматриваются в качестве архаичной группы, близкой к примитивным териодонтам. В Anteosauroidea автор различает только 2 семейства: Brithopodidae с подсемействами Brithopodinae и Anteosaurinae и Titanosuchidae с подсемействами Titanosuchinae и Tapinocephalinae. Таким образом, Д. Кинг попыталась покончить с разделением диноцефалов на преимущественно хищных титанозухов и вполне растительноядных тапиноцефалов.
У Д. Сигоньо-Расселл (1989 г.), в системе которой диноцефалы отсутствуют, из прежних диноцефалов в число примитивных териодонтов попадает семейство Eotitanosuchidae и частично также Biarmosuchidae. Однако многие исследователи продолжают рассматривать диноцефалов как промежуточную группу между сфенакодонтными пеликозаврами и аномодонтами.
Систему диноцефалов тщательно разрабатывал М.Ф. Ивахненко (2003, 2008 гг.), описавший ряд примитивных форм из России. Он признает самостоятельность диноцефалов, но считает эстемменозухов растительноядными потомками близких к горгонопсам эотериодонтов и даже относил эстемменозухов к подотряду горгонопсий. По мнению Л.П. Татаринова (2009 г.), эстемменозухов все-таки правильнее включать в состав диноцефалов и рассматривать как сестринскую по отношению к аномодонтам группу, ответвившуюся от примитивных хищных диноцефалов и находящуюся практически на уровне эотериодонтов. Помимо обычных подразделений диноцефалов (титанозухов и тапиноцефалов), М.Ф. Ивахненко (2008 г.) некоторое время выделял и третий, по его системе, самый примитивный инфраотряд — Eotitanosuchida, близкий по содержанию к установленному Э. Олсоном (1962 г.) инфраотряду Eotheriodontia.
М.Ф. Ивахненко также сделал важное для номенклатуры замечание о статусе распространенного ранее наименования Brithopodidae, которое он признал невалидным. Название данного семейства оказалось неудачным в силу неполноты остатков и неопределенного положения в ситеме рода, по которому оно было присвоено. С М.Ф. Ивахненко в дальнейшем согласился Л.П. Татаринов (2009 г.). По его мнению, валидным названием для аналогичной по объему группы диноцефалов, не включающей Titanosuchidae в понимании Д. Кинг (1988 г.), должно стать Titanophoneidae.
В разработке систематики диноцефалов необходимо также учитывать многие достижения Л. Бунстра (1952, 1954, 1964, 1965, 1968 гг. и др.).
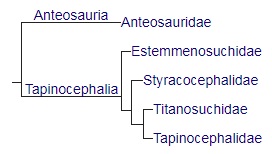
Кладограмма, иллюстрирующая современные взгляды на родственные связи диноцефалов (по Фрэзер-Кинг с соавт., 2019).
Здесь для отряда принят следующий состав:
подотряд †Титанозухиды (Titanosuchida);
подотряд †Тапиноцефалиды (Tapinocephalida).
Ссылки
Дейноцефалы (Википедия) перейти
Dinocephalia (Paleobiology Database) перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Ивахненко М. Ф. Тетраподы Восточно-Европейского плаката — позднепалеозойского территориально-природного комплекса. — Пермь, 2001
Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1 / Отв. ред. Ивахненко М. Ф. и Курочкин Е. Н. — М.: ГЕОС, 2008
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Протеро Д. Отпечатки жизни. 25 шагов эволюции и вся история планеты. — СПб.: Питер, 2016
Ромер А. Ш. Палеонтология позвоночных. — М. — Л.: Государственное научно-техническое издательство нефтяной и горно-топливной литературы, 1939
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГЕОС, 2009