Диноцераты — Dinocerata
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Клада Скротиферы (Scrotifera)
Клада Феревунгуляты (Fereuungulata)
Клада Эвунгуляты (Euungulata)
Клада Мезаксонии (Mesaxonia)
Отряд †Диноцераты (Dinocerata)
Диноцераты (Dinocerata Marsh, 1872) — отряд клады мезаксоний (Mesaxonia Marsh, 1884), происходящий из среднего палеоцена - среднего эоцена Северной Америки и позднего палеоцена - среднего эоцена Азии. Установлены 3 семейства с приблизительно 7 родами.
Разнообразие диноцератов (Dinocerata), слева направо: эобазилевс (Eobasileus cornutus), батиопсис (Bathyopsis fissidens), гобиатерий (Gobiatherium mirificum) и продинацер (Prodinoceras martyr). Автор — Д. Богданов.
История изучения
Первое описание остатков этих животных было сделано О.Ч. Маршем в 1871 г. под наименованием Titanotherium? anceps. Затем выходит серия работ О.Ч. Марша, Э.Д. Копа и Д. Лейди. Маршем в 1872 г. впервые применяется название Dinocerata для обозначения этого отряда, а в следующем, 1873 г., этот исследователь приводит общее описание группы, официально присвоив ей данное наименование. Оно образовано древнегреческими словами deinós — "ужасный" и kéras — "рог", что вместе переводится примерно как "страшнорогие". В этом же году Коп относит диноцератов к отряду хоботных в качестве особого подотряда. Саблевидные клыки этих животных он считал резцами. А.Г. Гаррод в работе 1873 г. рассматривал диноцератов как группу, относящуюся к парнопалым, какого мнения он придерживался и впоследствии (1881 г.).
В 1877 г. Dinocerata были включены Копом в качестве подотряда в отряд Amblypoda. Этой последней точки зрения длительное время придерживались очень многие исследователи. Например, Скотт в своей книге об истории млекопитающих Западного полушария (1937 г.) объединяет пантодонтов и диноцератов в один отряд. Безосновательность подобного объединения была впоследствии частично доказана Д.Г. Симпсоном (1929, 1945 гг.) и подробно разобрана К.К. Флеровым (1957 г.) на основании строения коренных зубов.
В 1886 г. появляется большая подробная монография Марша о диноцератах. Это весьма солидное сочинение содержит подробное описание скелета поздних американских диноцератов, относимых к семейству Uintatheriidae. В ней приводится их сравнение с другими млекопитающими, систематика, краткие геологические данные, общий взгляд на филогенетическое положение отряда. Функциональная морфология, в особенно экологии, совершенно не освещаются Маршем, так же как и всеми остальными учеными, за исключением Скотта (1913, 1937 гг.), который впервые дал более или менее серьезное и обоснованное морфо-экологически толкование различных особенностей строения.

Смонтированный скелет уинтатерия (Uintatherium anceps).
После работ Марша, Лейди, Копа и Скотта, которые описали ряд представителей поздних, среднеэоценовых американских диноцератов, относящихся к семейству Uintatheriidae, сравнительно долгий период, с 1890-х гг. до работ Центральноазиатской экспедиции Американского музея естественной истории, по этой группе не было сделано почти никаких крупных находок. Лишь в 1913 г. Осборн опубликовал описание черепа самого раннего на тот момент представителя уинтатериевых —
Bathyopsis. Обнаруженные названной экспедицией остатки диноцератов послужили основанием для ряда исследований и вновь вызвали интерес к изучению этой группы.
В 1929 г. Метью, Грейнджер и Симпсон по весьма фрагментарным остаткам описали древнейшего известного представителя диноцератов — Prodinoceras из позднепалеоценовой формации Гашато в Монголии. Несколько позднее, в 1930 г., в среднеэоценовых отложениях Ирдын-Манга во Внутренней Монголии той же экспедицией были найдены черепа и части скелета необычного диноцерата Gobiatherium (Осборн и Грейнджер, 1932 г.). К этому же времени относится ряд находок Prodinoceras в позднем палеоцене Северной Америки, откуда были описаны синонимичные ему Probathyopsis (Симпсон, 1929 г.; Джепсен, 1930 г.) и Bathyopsoides (Паттерсон, 1939 г.).
Значительно дополнили знания об этих замечательных животных работы Монгольской палеонтологической экспедиции Академии наук СССР 1946-1945 гг. В Нэмэгэтинской котловине, расположенной в Южной Гоби, в районе хребта Цаган-Ула, у источников Наран-Булак и Улан-Булак было открыто кладбище раннепалеогеновых млекопитающих, среди которых подавляющая часть остатков принадлежит примитивным диноцератам продиноцерам (младший синоним — Mongolotherium). Посвященная анализу этих находок работа К.К. Флерова «Диноцераты Монголии» (1957 г.) не утратила своей высокой научной ценности и в наши дни.
Ископаемые остатки диноцератов (Dinocerata): вверху слева — часть черепа эобазилевса (Eobasileus cornutus), вверху справа — неполный череп гобиатерия (Gobiatherium mirificum), внизу — черепа, нижние челюсти и фрагменты скелета продиноцера (Prodinoceras martyr). Палеонтологический музей им. Ю.А. Орлова РАН, Москва.
Происхождение и родственные связи
В монофилетическом, общем происхождении диноцератов не существует никаких сомнений — об этом ясно свидетельствуют многочисленные и своеобразные особенности их строения. Что же касается их родственных связей, то они все еще остаются невыясненными.
Длительное время диноцератов сближали с Pantodonta и помещали обе группы в отряд Amblypoda. При этом указывалось, что у них имеются общие черты в строении зубного ряда, предплюсны и ряда других элементов посткраниального скелета. В дальнейшем, однако, было установлено, что некоторое морфологическое подобие (конвергенция) диноцератов и пантодонтов объясняется их сходным образом жизни, и амблиподы были расформированы на два самостоятельных отряда. В настоящее время допускается их независимое происхождение и самостоятельное развитие. Видимо, диноцераты произошли от более древних и архаических предков, чем пантодонты, у которых даже древнейшие формы имеют зубы более сложного строения.
Впоследствии диноцератов стали считать рано уклонившейся ветвью настоящих копытных — Ungulata. Но даже в составе этой обширной группы положение диноцератов неясно. Порой их предками считают Arctocyonia, некогда определяемых в ныне спорный таксон Condylarthra. В одно время диноцератов также пытались сблизить с надотрядом Paenungulata, включающим хоботных, сирен, даманов и др. Сходство со слонами в строении скелета поздних Uintatheriidae настолько велико, что некоторые ученые даже относили их к хоботным. Само собой разумеется, подобная классификация не имела под собой никаких действительных оснований и была совершенно ошибочна.

Реконструкция внешнего облика эобазилевсов (Eobasileus cornutus, = Loxolophodon cornutus), отображающая представление Э. Копа о принадлежности диноцератов к хоботным. Автор — Э. Шеппард, 1873 г.
Некоторые авторы указывают на сходство диноцератов с Pyrotheria и Xenungulata, полагая, что данные таксоны можно объединить в группу Uintatheriamorpha. Отмечается также сходство в строении зубной системы уинтатериеобразных не с прочими копытными, а с Pseudictopidae из Anagaloidea — примитивными мелкими млекопитающими из Восточной Азии, родственными зайцеобразным и грызунам. Псевдиктоповые напоминают диноцератов типом моляризации задних предкоренных, V-образными лофами на верхних зубах и мезиодистально сжатыми тригонидами с широкими протолофидами (металофидами) на нижних коренных. Однако гипотеза подобного родства не получила достаточной поддержки среди специалистов. Ими указывается, что специализированный, сложенный по кроличьему типу скелет псевдиктоповых имеет мало общего с массивным, громоздким скелетом первых диноцератов, а сходство некоторых деталей зубного строения обеих групп поверхностно.
Иногда указывают на схожее строение III-х коренных зубов диноцератов и Carodnia — продвинутого ксенунгулята из позднего палеоцена Южной Америки. Однако, делать на этом основании выводы о эволюционной близости обеих групп явно преждевременно, поскольку морфологическое сходство остальных зубов далеко не так выражено. Кроме того, зубной аппарат более примитивного ксенунгулята Etayoa напоминает таковой диноцератов в гораздо меньшей степени. Хотя вопрос пока не решен окончательно, очевидно, правы те авторы, которые считают сходство в строении коренных зубов диноцератов и ксенунгулят чисто конвергентным. Существует также предположение о родстве с Zhelestida, группой эвтериевых млекопитающих из мелового периода.
Ближайшими ныне живущими родственниками диноцератов, по всей видимости, являются непарнопалые (Perissodactyla). Для обеих групп характерно мезаксонное положение ступни (ее главная ось проходит через средний, III-й луч) и отсутствие глубокой борозды на таранной кости для сочленения с голенью, что часто встречается, например, у афротериев (Afrotheria). В любом случае, диноцераты представляли собой специализированную, относительно недолго просуществовавшую боковую ветвь.
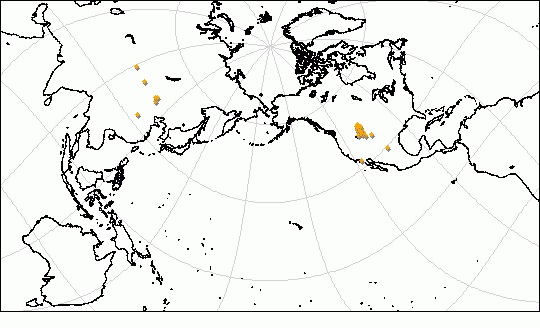
Возраст и распространение
Наиболее вероятно, отряд имеет североамериканское происхождение. Сдесь его представители известны начиная со среднего палеоцена (I-я половина тиффанского века). В позднем палеоцене (низы гашатского яруса) они появляются и в Азии. В Северной Америке эти животные обитали до конца I-й половины среднего эоцена (конец уинтского века), а в Азии их специализированные представители задерживаются до конца среднего эоцена (бартонский ярус).
Основные местонахождения ископаемых остатков диноцератов (Dinocerata).
Состав
В зависимости от варианта классификации, в отряд включают 3, 2 или только 1 семейство. Однако следует отметить, что подобный редукционизм напрямую вытекает из подхода к изучению диноцератов американскими специалистами. Большинство их работ по данной группе характеризуются чрезмерной лаконичностью и практически полным отсутствием функциональных и экологических трактовок рассматриваемого материала. Иногда лишь вкратце приводятся некоторые филогенетические соображения. Несомненно, результатом этого было и остается то, что все диноцераты нередко объединяются в единственное семейство Uintatheriidae, без сколько-нибудь серьезной попытки проанализировать в целом их филогению и систематику. Здесь для отряда принимается следующий состав.
Продиноцератовые (Prodinoceratidae Flerow, 1952) — самые древние и примитивные диноцераты. Представлены единственным родом Prodinoceras, имеющим много различных синонимов (Mongolotherium, Probathyopsis и т. д.). Род представлен как в Азии (Монголия, Китай), так и в Северной Америке (запад США, преимущественно Вайоминг), где его многочисленные виды обитали с позднего палеоцена по ранний эоцен.
Уинтатериевые (Uintatheriidae Flower, 1876) — более продвинутые ранне- и среднэоценовые диноцераты. Являются крупнейшими представителями отряда. Включают около 4 родов: Bathyopsis, Uintatherium, Tetheopsis и Eobasileus с множеством синонимов. Долгое время считалось, что семейство эндемично для Северной Америки (Вайоминг, Колорадо, Юта, Калифорния, Техас), но затем их обнаружили и в Китае.
Гобиатериевые (Gobiatheriidae Flerow, 1952) — высокоспециализированные диноцераты из раннего и среднего эоцена Азии (Китай, Казахстан, Киргизия). Представлены единственным родом Gobiatherium. Иногда его относят к Uintatheriidae, выделяя в их составе в собственное подсемейство. Пока не ясно, произошло ли группа от американских иммигрантов, или имеет азиатские корни.
Строение и функции
Все известные диноцераты — животные средней или значительной величины. Череп длиной от 30 см у ранних форм (
Prodinoceras) до 98 см у поздних (
Tetheopsis). Общая длина варьирует в пределах 2-4,5 м, высота в холке — 0,7-2,1 м, вес оценивается в 150-4000 кг.

Смонтированный скелет, его прорисовка и реконструкция внешнего облика уинтатерия (Uintatherium anceps). Автор реконструкции — Д. Богданов.
Черепа ранних и поздних представителей отряда обладают различными пропорциями. Длина лицевого отдела может быть как меньше, так и почти равной длине мозгового. Глазница позади без костной дуги, широко открыта. Верхние заглазничные отростки очень малы, нижних нет. Скуловые дуги раздвинуты в стороны в разной степени. Слуховой пузырь отсутствует, барабанная кость не полностью окостеневшая. У примитивного рода Prodinoceras чрезвычайно развит сагиттальный гребень — сильнее, чем у каких-либо других млекопитающих.
Начиная с раннего эоцена Uintatheriidae приобретают парные выросты на верхней стороне черепа. У раннего уинтатериида
Bathyopsis, морфологически и стратиграфически занимающего промежуточное положение между Prodinoceratidae и более продвинутыми представителями собственного семейства, наряду с исчезновением сагиттального гребня на черепе впервые появляются небольшие костные бугорки. У более поздних, ранне- и среднеэоценовых родов, они превращающиеся в массивные рогообразные выросты числом от 1 до 3 пар, расположенные соответственно на носовых, верхнечелюстных и теменных костях. Кроме того, дополнительная пара небольших фронтальных выступов может располагаться чуть выше передней части глазниц.

Череп с нижней челюстью уинтатерия (Uintatherium anceps), вписанный в телесные очертания.
Передняя, носовая, пара выростов у Uintatheriidae мала и, возможно, была опорой для состоящих их кератина рогов, похожих на рога парнорогих вымерших носорогов рода Diceratherium, остальные две пары гораздо больше. Судя по структуре поверхности, верхнечелюстные выросты, вероятно, не имели роговых чехлов и не служили основанием для спадающих рогов, а были покрыты кожей, как оссиконы Giraffa. Форма теменных выростов также не позволяет допустить наличие рогового чехла. Если же они все-таки и были покрыты роговыми футлярами, то они должны были периодически сбрасываться и вновь нарастать, как у Antilocapra. Эта третья пара выростов, отходящих от теменных костей, массивная, расширяющаяся булавовидно к концу. Поперечный затылочный гребень очень высок; от него, справа и слева, отходят вперед теменные гребни, несущие теменные "рога". Гребни эти проходят над глазницами, заходя дальше них вперед. Крыша черепа представляет собой плоскую или даже вогнутую широкую поверхность, открытую спереди. Среди других млекопитающих подобная впадина имеется лишь у некоторых Brontotheriidae.
В сравнении с размерами черепа и всего тела мозговая полость очень невелика и занимает лишь задне-нижнюю часть черепа. Слабо дифференцированный головной мозг всех диноцератов крайне мал. По своим относительным размерам он значительно меньше, чем у ныне живущих копытных и лишь немного превышает мозг рептилий, что свидетельствует о чрезвычайно низком интеллектуальном уровне этих животных. Крыша черепа и слепки чрезвычайно толсты, но облегчены тем, что имеют губчатую структуру и, кроме того, заключают в себе систему синусов. Однако как синусы, тая и губчатость кости выражены у диноцератов слабее, чем у других крупных млекопитающих, как, например, у слонов, носорогов и бронтотериев, вооруженных тяжелыми бивнями и рогами.

Относительные размеры головного мозга, слева направо: лошадь Equus, бронтотерий Megacerops, пантодонт Coryphodon, диноцераты Uintatherium и Tetheopsis.
Предчелюстные кости у Prodinoceratidae хорошо развиты и служат опорой для верхних резцов. У Uintatheriidae и Gobiatheriidae в связи с утратой резцов они становятся слабыми, утончаясь к концам, и не соприкасаются спереди друг с другом. Такие лишенные зубов предчелюстные кости, несомненно, несли жесткие небные складки, противостоящие нижним резцам при срезании и откусывании листьев и другой мягкой растительности. Вполне вероятно, для ее захватывания имелась подвижная верхняя губа. Носовые кости длинны и массивны; они нависают над предчелюстными спереди, что вместе с формой и положением переднего носового отверстия указывает на отсутствие хобота. У Gobiatherium носовые кости аркообразно вздуты.
Нижняя челюсть с высокой восходящей ветвью и длинным венечным отростком, угловой отросток в виде широкой лопасти. Суставный валик у ранних форм располагается почти на уровне коренных зубов, у более поздних — несколько выше. Нижнечелюстной симфиз плотно сросшийся. Вблизи симфиза у большинства диноцератов присутствуют более или менее выраженные костные лопасти, выступающие вниз и в стороны и предохраняющие ложащиеся в них увеличенные верхние клыки. Из всех известных диноцератов эти лопасти отсутствуют только у одного Gobiatherium, не имеющего верхних клыков, обладающего суженным, очень низким и уплощенным черепом без выраженных рогообразных выростов на верхней части, но в то же время с высоко поднятыми носовыми костями и расширенным симфизом на невысокой нижней челюсти. Соответственно развитию клыков у самцов большинства диноцератов лопасти выражены сильнее, чем у самок, причем у самых поздних родов самки вообще их теряют. Древний род Prodinoceras имеет самок с довольно длинными верхними клыками, почему и лопасти на нижних челюстях у них выражены совершенно отчетливо, хотя всегда значительно слабее, чем у самцов. Это указывает на то, что половой диморфизм у архаичных диноцератов был слабо выражен, и клыки несли иную функцию, чем впоследствии, когда они стали настоящим "турнирным" оружием. Параллельно с ослаблением клыков у самок в ветви Uintatheriidae шло развитие у самцов рогообразных выростов. Сочленовные бугорки нижней челюсти расположены на ножках и выдвинуты назад, а не вверх, так как диноцераты, подобно саблезубым кошкам, обладали способностью чрезвычайно широко раскрывать рот. Подобное строение нижней челюсти вместе с чрезвычайно сильным развитием гребней на черепе, увеличивающих площадь мест прикрепления мускулатуры, смыкающей челюсти, свидетельствует о том, что эти животные проделывали клыками значительную работу.
Портретные реконструкции диноцератов (Dinocerata). Автор — Г. Угуэто.
По своему строению зубы различных диноцератов весьма сходны и в целом характеризуются значительной примитивностью. Зубная формула представителей отряда: I0-3/i3, С0-1/c1, Р3-4/p3-4, М3/m3. У ранних форм присутствует почти полный набор зубов (кроме Р1). Prodinoceras обладает хорошо развитыми верхними резцами, у более продвинутых форм они исчезают и вся зубная система приобретает значительное сходство с зубной системой жвачных. Нижние резцы большие, у Prodinoceras их коронки уплощены, сужены к вершине, с зазубренным волнистым краем. В дальнейшем, у Uintatheriidae и Gobiatheriidae, коронки нижних резцов постепенно расширяются от i1 к i3, приобретают несколько вершинок на своей режущей кромке, конический острый конец спереди и длинную пятку позади, которая иногда несет на себе маленькую вершинку, а у поздних родов (например, у Eobasileus и Gobiatherium) получают дополнительную наружную лопасть.
Верхние клыки обычно сильно развитые, саблевидно изогнуты назад, реже прямые, могут быть значительно удлинены. Они уплощены со сторон, с острыми гребнями вдоль переднего и заднего края, причем задний гребень слегка зазубренный, но не образует такой правильной пильчатости как у части горгоноповых, хищных динозавров и саблезубых кошек. Передний гребень у ранних родов слабо выражен. Клыки постепенно конически суживаются к заостренной вершине или иногда несколько расширяются и на форме напоминают наконечник копья. Как уже упоминалось, особенно большого размера верхние клыки достигают у взрослых самцов, у самок и молодых самцов они значительно меньше (то же верно и в отношении костных лопастей на нижней челюсти). Верхние клыки у самок всех родов значительно короче и тоньше, чем у самцов; это, несомненно, доказывает, что они служили у последних оружием для борьбы между собой, подобно тому, как это имеет место у парнопалых. Нижние клыки у палеоценовых и раннеэоценовых диноцератов были велики, почти трехгранные в сечении, конические. Они имели резко выраженные острый передний и задний гребни, наружную сторону округленную, а внутреннюю плоскую. У более поздних родов нижние клыки уменьшаются и приобретают форму и функцию резцов, вследствие чего зубная формула еще более приближается к жвачным, в особенности у
Gobiatherium с утраченными верхними клыками. Исчезновение верхних резцов шло в процессе развития диноцератов параллельно с уменьшением нижних клыков и приобретением ими резцеобразной формы, что свидетельствует о смене пищевого режима и способа добывания пищи.

Расположение бугров на левом верхнем (А) и левом нижнем (Б) коренных зубах уинтатерия (
Uintatherium anceps): cing ant — передний цингулюм; cing post — задний цингулюм; end — энтоконид; endсr — энтоконидная криста; hld — гипоконулид; hldcr — гипоконулидная криста; һу — гипокон; hyd — гипоконид; mе — метакон; med — метаконид; ml — металоф; msd — метастилид; ра — паракон; pad — параконид; pl — протолоф; рг — протокон; prd — протоконид.
Предкоренные диноцератов сильно моляризованы; коренные с низкими коронками, лофодонтные, увеличивающиеся в заднем направлении. На жевательной поверхности верхних щечных зубов (Р3-М3) имеются поперечные эмалевые гребни, представляющие собой протолоф (паралоф) и металоф, отходящие от языковых поверхностей паракона и метакона и сходящиеся в протоконе, образуя таким образом на каждом зубе V-образную, направленную внутрь фигуру. Присутствуют широкие передние и задние цингулюмы, зачастую непрерывающиеся по всей языковой границе. На нижних щечных зубах, пока они не стерты, гребни покрыты мелкими бугорками; особенно выделяются высокие металофиды (протолофиды), располагающиеся между протоконидом и метаконидом. Гораздо ниже гребешок косого кристида, простирающийся от гипоконида к выраженному метастилиду, что вообще очень характерно для нижних зубов различных представителей отряда. Параконид и паралофид рудиментарны либо полностью отсутствуют. За несколькими известными исключениями, зубы диноцератов совершенно не похожи на зубы других млекопитающих, поэтому гомология их частей до сих пор очень спорна. Поримечательно, что у позднейших родов щечные зубы по отношению к общим размерам черепа очень малы. Это объясняется тем, что они незначительно и непропорционально увеличивались в размерах по сравнению с общим ростом животных в процессе истории развития группы. Поэтому зубы у гигантских форм лишь немногим крупнее, чем у мелких (приблизительно на одну треть), в то время как черепа
Uintatherium и
Eobasileus не менее чем вдвое больше, чем у
Prodinoceras.
Большой интерес представляет конвергенция в форме черепа диноцератов с некоторыми парнопалыми — например, родом Protoceras. Сходство не ограничивается саблеобразностью клыков и отсутствием верхних резцов, а идет значительно дальше. Череп Protoceras, подобно черепу Uintatheriidae, несет на верхней стороне рогообразные выросты — одну пару на теменных и одну на лобных костях; кроме того, пара плоских выростов образована верхнечелюстными костями и расположена на носу (над клыками и предкоренными); меньших размеров плоские выросты развиты на лобных костях над орбитами. Подобно диноцератам, у Protoceras все это вооружение, как и саблезубость клыков, значительно выражено у самцов и очень слабо у самок. Интересно отметить, что коренные зубы Protoceratidae относительно мелкие. Несомненно, в данном случае мы имеем пример не только внешнего, но и функционального сходства.
Посткраниальный скелет всех диноцератов массивный. Шея относительно длинная. Конечности мощные, короткие или умеренной длины, передняя их пара обладает более или менее выраженной способностью прогибаться в локтевых суставах. Интермедиальные элементы конечностей короче, чем проксимальные, метаподии обычно короткие и крепкие. Локтевая кость свободная, развита сильнее лучевой. В запястье ладьевидная и полулунная кости раздельны, свободная центральная отсутствует. Малая берцовая кость цельная, не сращена с большой; в заплюсне I-я и III-я клиновидные кости сращены или раздельны. Кисть и стопа короткие, но широкие. На конечностях по 5 коротких пальцев, конечные фаланги — в виде округлых наростов, несут в разной степени расширенные копыта. Поздний уклонившийся род
Gobiatherium выделяется относительно удлиненными стройными метаподиями. Как вообще у крупных и массивных млекопитающих, длинные кости диноцератов не имели костномозговой полости, а были заполнены губчатой костной массой. Подобная структура значительно увеличивает прочность без увеличения диаметра кости.

Прорисовки скелета кисти (А) и стопы (Б) уинтатерия (Uintatherium anceps).
У раннего рода Prodinoceras, имеющего относительно небольшие размеры тела, посткраниальный скелет еще весьма архаичен. У этих животных было удлиненное туловище, и короткие, полупальцеходящие передние и стопоходящие задние конечности. От продвинутых диноцератов их также отличает наличие ключицы, III-го вертлуга на бедренной кости, мелкая рифленая таранная кость с короткой шейкой, сращенные клиновидные кости, относительно узкий таз и длинный, хорошо развитый хвост, составляющий около 1/3 всей длины позвоночника. Более поздние формы (Uintatherium, Eobasileus и др.) представляют собой тяжеловесных носорого- или даже слонообразных животных, достигавших более 2 м в холке, с колоннообразными пальцеходящими передними и задними конечностями. Подобно таковым слонов, они вполне могли иметь амортизирующую эластическую подушку. Бедро этих диноцератов сравнительно длиннее голени, с уплощенным телом без III-го бедренного вертлуга, элементы предплюсны широкие и сжаты в переднезаднем направлении, короткая таранная кость обладает плоским блоком и лишена шейки, клиновидные кости раздельны, таз широкий, а хвост имеет небольшую длину. Массивное туловище имело большой объем, о чем можно судить по длине и изгибу ребер. Относительно наличия волосяного покрова сказать что-либо определенное трудно. Не исключено, что он сохранялся не только у архаичных, но и у поздних форм.
Образ жизни
Об образе жизни диноцератов судить сложно. Несмотря на увеличенные клыки, лофодонтные щечные зубы свидетельствуют о растительноядном рационе. Наиболее древние диноцераты, однако, вполне могли быть и всеядными. Зубочелюстной аппарат более поздних представителей уже относительно специализирован. Низкие, мало стирающиеся с возрастом коронки щечных зубов указывают на то, что животные не могли питаться грубой пищей и потребляли только мягкую растительность. При закрытии челюстей верхние и нижние зубы смыкались в крепкий замок, исключающий выраженные переднезадние и боковые движения челюстей. Таким образом, пища в основном не перетиралась, а просто раздавливалась силой челюстных мышц. Саблевидные клыки диноцераты могли использовать при добывании корма, а также как турнирное оружие.

Уинтатерии (Uintatherium anceps) в естественном окружении. Автор — З. Буриан.
Ранние диноцераты, Prodinoceratidae, оставались еще сравнительно небольшими животными. Строение их задних конечностей указывает на то, что звери вполне могли на них подниматься, а кисти в небольшой степени сохранили способность к манипуляциям. Продиноцератовые питались сочной растительной пищей, возможно, иногда разнообразя свой рацион мелкой добычей или падалью. Видимо, они обитали в лесных биотопах, не слишком близко к водоемам. В дальнейшем диноцераты стали крупными, строго растительноядными животными с массивным туловищем и непропорционально объемистым тазом, что, вероятно, было связано со значительной продолжительностью кишечника. Возможно, в нем у поздних представителей группы, подобно лошадям и сиренам, происходил процесс ферментации поглощенной растительной пищи, компенсирующий ее достаточно грубую обработку в ротовой полости.
По своей морфологии, пропорциям и размерам тела диноцераты напоминают самых различных млекопитающих — носорогов, гиппопотамов, слонов, сирен, поэтому понять, какой именно образ жизни они вели, трудно. Возможно, экологически поздние представители отряда представляли собой нечто среднее между вышеперечисленными животными. Бочкообразная грудная клетка и отсутствие воздушных полостей в пахиостозно утолщенных длинных костях конечностей могут свидетельствовать о более или менее выраженной амфибиальности. Тяжелые кости компенсируют плавучесть тела и характерны для целого ряда наземных позвоночных, вторично адаптировавшихся к жизни в водной среде. Вероятно, Uintatheriidae были привязаны к влажным биотопам, однако в какой степени, сказать трудно. Gobiatheriidae, очевидно, вели полуводный образ жизни, добывая пищу преимущественно в воде.

Сражающиеся самцы уинтатерия (Uintatherium anceps). Автор — В. Симеоновски.
Вымирание
Диноцераты являлись одними из первых по-настоящему больших млекопитающих. Очевидно, таких размеров они сумели достичь потому, что после вымирания динозавров ниша крупных растительноядных животных была свободна, и серьезных конкурентов у них отсутствовали. В ходе своей эволюции диноцераты не смогли дать значительного разнообразия форм, а количество их родов было ограниченным. Несмотря на это, вплоть до полного исчезновения в среднем эоцене они практически всегда заступали в своих экосистемах места наиболее крупных фитофагов. Мелкие эоценовые диноцераты почти не известны. Вероятно, им сложно было конкурировать с новыми, более прогрессивными группами копытных.
К угасанию диноцератов привело стечение ряда неблагоприятных обстоятельств. По сути своей они являлись животными архаичными, обладали консервативным планом строения и невысокими адаптивными способностями. Пока климат был теплым и влажным, эти звери процветали, расселялись и все более увеличивались в размерах. Однако принципиальных изменений в их морфологии не происходило, а громоздким телом продолжал управлять маленький и слаборазвитый головной мозг. Когда природная обстановка стала меняться, постепенно начали исчезать не сумевшие приспособиться к новым условиям группы. Среди них оказались и диноцераты. Не исключено, что их исчезновению способствовала и конкуренция с первыми крупными непарнопалыми из числа бронтотериев и аминодонтов, появившимися к середине эоценовой эпохи. Как бы то ни было, древний отряд вымер, не оставив потомков.
Ссылки
Диноцераты (Большая российская энциклопедия)
перейти Dinocerata (Paleobiology Database)
перейти
Литература
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 3 — М.: Мир, 1993
Молош А. Альтернативные звери раннего палеогена. В мире животных 2, 2012
Основы палеонтологии. В 15 т. Т. 13: Млекопитающие / Гл. ред. Орлов Ю. А. — М.: Государственное научно-техн. изд-во литературы по геологии и охране недр, 1962
Павлова М. В. Палеозоология. Ч. 2. Позвоночные. — М.-Л.: Государственное изд-во, 1929
Ромер А. Ш. Палеонтология позвоночных. — М. — Л.: Государственное научно-техн. изд-во нефтяной и горно-топливной литературы, 1939
Россолимо О. Л., Павлинов И. Я., Крускоп С. В., Лисовский А. А., Спасская Н. Н., Борисенко А. В., Панютина А. А. Разнообразие млекопитающих. Ч. 1-3. — М.: Изд-во КМК, 2004
Тернер А., Антон М. Большая энциклопедия доисторических животных — М.: Оникс, 2006
Флеров К. К. Диноцераты Монголии. Труды палеонтологического института. Том 67. М.: Академия наук СССР, 1957
Evolution of Tertiary Mammals of North America. Vol. 1: Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals / Ed. by Janis C. M., Scott K. M. & Jacobs L. L. — Cambridge: Cambridge University Press, 1998
Kemp T. S. The Origin and Evolution of Mammals. — Oxford: Oxford University Press, 2005
Marsh O. C. Dinocerata. A monograph of an extinct order of gigantic mammals. — Washington: United States Geological Survey, 1886
Rose K. D. The Beginning of the Age of Mammals. — Oxford: Johns Hopkins University Press, 2006
Wheeler W. H. Revision of the uintatheres. Peabody Museum of Natural History Bulletin 14, 1961