Размеры варьируют от мелких до очень крупных.
Верхняя часть аддукторной полости, расположенная над областью crista parotica, значительно превышает по высоте нижнюю. Чешуйчатая кость не имеет шовного латерального контакта с квадратно-скуловой костью. Квадрато-квадратно-скуловой комплекс расположен в специальном углублении затылочной пластины чешуйчатой кости (стрептостилия).
Характерно развитие разных типов зубного аппарата с первыми попытками создать вторичное мягкое небо, позволяющее одновременно есть и дышать.
Открытие в средней перми Урала примитивных хищных терапсид, более плезиоморфных, чем типичные териодонты, вызвало дискуссии о положении этих терапсид в системе. В большинстве случаев они имеют неясные связи с предковыми диноцефалами, у их потомков иногда прослеживаются связи с горгонопсами. Л.П. Татаринов (2009 г.) объединяет эти формы с близкими к ним формами из других регионов под условным названием "фтинозухии", оговаривая при этом, что эта группа разнородная и, скорее всего, сборная.
А. Ромер (1961 г.) установил для родов Phthinosuchus и Phthinosaurus из средней перми Башкортостана особый инфраотряд Phthinosuchia, в котором он видел вероятных предков териодонтов и аномодонтов. В отличие от собственно териодонтов, фтинозухии не имели развитого венечного отростка, который у них практически не выступает над верхним краем нижней челюсти, височная область черепа у них короткая, число заклыковых зубов относительно велико — не менее 10. Ранее этих терапсид относили то к диноцефалам (Сили, 1894 г.), то к пеликозаврам (И.А. Ефремов, 1940 г.), то, наконец, к горгонопсам в качестве особого семейства последних (И.А. Ефремов, 1954 г.). Позднее Э. Олсон (1962 г.) установил инфраотряд Eotheriodontia, в котором он объединил всех примитивных хищных терапсид: помимо фтинозухид также и эотитанозуха и биармозухид из открытой перед тем среднепермской фауны Очера в Пермской области (П.К. Чудинов, 1960 г.), а также ранее известных бритоподид вместе с описанными лишь по незначительным фрагментам проблематичными хищными терапсидами из ранннепермской формации Сан-Анджело в США.
Позднее Л. Бунстра (1963 г.) установил подотряд Eotitanosuchia, включающий только фтинозухид и эотитанозухид. Л.П. Татаринов (1974 г.) выделил фтинозавра в новое семейство эотериодонтов, а чуть позднее тот же Л.П. Татаринов (1976 г.) присоединил фтинозухий, возведенных в ранг отдельного подотряда, к горгонопсам, принимаемым в ранге отряда; это осталось, по всей вероятности, только предположением. Фтинозухий объединяли в одну группу с горгонопсами также Т. Кемп (1982 г.) и А. Бринк (1982, 1986 гг.), но первый автор предпочитал называть объединенную группу, принимаемую им в ранге подотряда, Eotheriodontia. Эотериодонтов Т. Кемп разделил на 2 инфраотряда — Eotitanosuchia (с семействами Phthinosuchidae и Eotitanosuchidae) и Gorgonopsia (с семействами Ictidorhinidae и Gorgonopsidae).
Д. Сигоньо-Расселл (1989 г.), однако, не приняла объединения горгонопсов с фтинозухиями и другими эотериодонтами, а, наоборот, выделила из состава горгонопсов морфологически более примитивные семейства Hipposauridae, Ictidorhinidae и Burnetiidae в инфраотряд Biarmosuchia. Все эти семейства, в отличие от настоящих горгонопсов, не имеют в нижней челюсти возвышающегося над supraangulare венечного отростка. Д. Сигоньо-Расселл (1989 г.) назвала биармозухов новым подотрядом, но в действительности эта группа была выделена ранее (Хопсон и Бархьюзен, 1986 г.; Роу, 1986, 1988 гг.). Остальных эотериодонтов Д. Сигоньо-Расселл разделила на 2 самостоятельных инфраотряда: Phthinosuchia (с семействами Phthinosuchidae и Phthinosauridae) и Eotitanosuchia (семейство Eotitanosuchidae). Напомним, что М.Ф. Ивахненко (2008 г.) считает роды Biarmosuchus и Eotitanosuchus синонимами.
М.Ф. Ивахненко и др. (1997 г.) сохранили самостоятельность горгонопсов, включаемых ими в состав териодонтов, но резко расширили объем эотериодонтов, принимаемых ими в ранге надотряда. Эотериодонтов они разделили на 2 отряда: Phthinosuchia и Niaftasuchia. Няфтазухии — отряд, описанный по единственному роду Niaftasuchus из средней перми Архангельской области. Первоначально няфтазух по признаку отсутствия клыков и листовидности коронок заклыковых зубов был отнесен М.Ф. Ивахненко (1995 г.) к примитивным тапиноцефалам. Но зачаточное захождение челюстных аддукторов в заглазничную область оказалось единственным признаком, явно отличающим няфтазуха от типичных тапиноцефалов. Выявленное позднее наличие межтеменной кости указывает на возможную близость няфтазуха к примитивным аномодонтам.
М.Ф. Ивахненко и др. (1997 г.) кардинально пересмотрели объем отряда Phthinosuchia. Помимо собственно фтинозухий и эотитанозухий, авторы включили в отряд аберрантное семейство Burnetiidae, эстемменозухов (ропалодонтид) и род Phthinosaurus. Бурнетиид рассматривают как семейство горгонопсов (Ватсон и Ромер (1956 г.), Ромер (1956 г.), Эйлер (1961 г.), Вьюшков (1964 г.), Татаринов (1974 г.)), но прежде многие исследователи относили их к диноцефалам (Хаутон (1924 г.), Хюне (1948, 1956 гг.). Р. Брум (Broom, 1923 г.) выделял их, вместе с диноцефалом Styracocephalus, в особый подотряд Burnetiamorpha, более близкий к горгонопсам, чем к диноцефалам, а Д. Сигоньо (1970 г.) одно время рассматривала их как Theriodontia incertae sedis.
Эстемменозухов, или ропалодонтид, относимых ранее к примитивным диноцефалам (Чудинов (1960, 1964, 1983 гг.), Кемп (1982 г.), Хопсон и Бархьюзен (1986 г.), Кинг (1988 г.), Хопсон (1991 г.)), М.Ф. Ивахненко и др. (1997 г.) считают эотериодонтами, вторично перешедшими к растительноядности. У эстемменозухов нет ни следа разрастания наружного челюстного аддуктора (m. adductor mandibulae externus) на заглазничную кость, что столь характерно для настоящих диноцефалов (Бунстра (1963 г.), Бархьюзен (1973, 1975, 1976 гг.), Ивахненко (1995 г.)). Следует все же отметить, что в меньшей степени, чем у диноцефалов, разрастание нижнечелюстных аддукторов свойственно также биармозухидам и эотитанозухидам (Хопсон и Бархьюзен (1986 г.), Ивахненко (1999 г.)), так что специфичность этого признака нельзя переоценивать.
Phthinosaurus отнесен М.Ф. Ивахненко и др. (1997 г.) к Phthinosuchia на основании треугольной формы надугловой кости, утолщенных в сечении клыков с гранями на поверхности и признаков листовидности задних заклыковых зубов, что, по мнению Л.П. Татаринова (1998, 2009 гг.), не вполне убедительно. Этот автор отмечает, что граненые в сечении клыки имеют и некоторые хищные тероцефалы, так что сам по себе этот признак не свидетельствует о растительноядности. Однако перед этим Л.П. Татаринов неудачно сравнил задний заклыковый зуб фтинозавра с примитивно триконодонтными зубами тероцефалов; в действительности этот зуб имеет грубую пильчатость по краям коронки, что придает ему сходство как с ропалодонтными зубами, так и с зубами аберрантного горгонопида Rubidgina.
Позднее М.Ф. Ивахненко (2000 г.) объединил горгонопсов с эотериодонтами в один отряд, разделив его на растительноядных ропалодонтов (эстемменозухов вместе с фтинозавром) и хищных горгонопсов; обе группы приняты им в ранге подотрядов. Эстемменозухов, или ропалодонтов, М.Ф. Ивахненко рассматривал в качестве растительноядных эотериодонтов, обладающих бивнеобразными округлыми в сечении клыками, короткими лепестковидными заклыковыми зубами и глубоким носоглоточным проходом. Для ропалодонтов характерны также пахиостозные разрастания покровных костей черепа, в меньшей степени выраженные у рубиджеид, отнесенных к настоящим горгонопсам, а в большей степени — у бурнетиид. По строению височного окна с расширенной заглазничной дугой и опущенной скуловой костью эстемменозухи сходны с рубиджеидами, с которыми они, по мнению М.Ф. Ивахненко, может быть, относительно близки по родству. В дальнейшем М.Ф. Ивахненко (2001 г.) включил ропалодонтов (=эстемменозухов) в отряд горгонопсов, подразделенных на 2 подотряда: ропалодонтов, к которым условно отнесены и микроураниоидеи, и собственно горгонопсов.
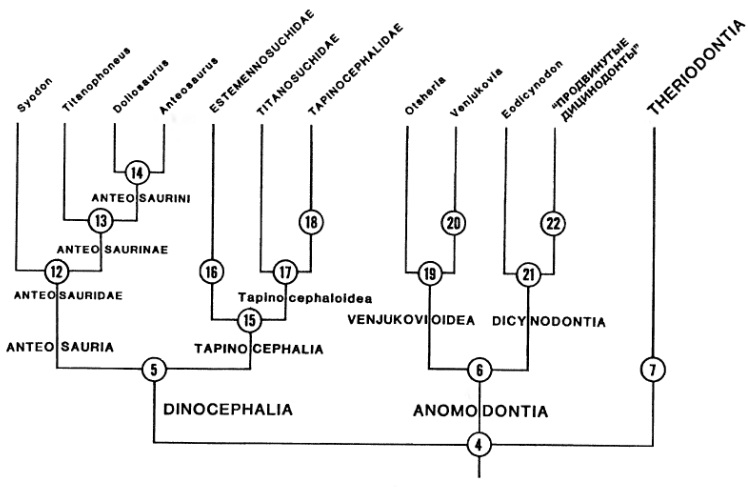
В одной из следущих работ М.Ф. Ивахненко (2003 г.) объединил диноцефалов в традиционном значении с горгонопсами и аномодонтами в инфракласс Eotherapsida, выделив в последнем надотряд Dinomorpha с отрядами Gorgodontia и Anomodontia. В этой работе М.Ф. Ивахненко в корне пересмотрел объем и систему отряда Gorgodontia, который подразделил на подотряды Dinocephalia и Gorgonopia. Подотряд Dinocephalia, близкий по объему к классическим титанозухидным диноцефалам, М.Ф. Ивахненко разделил на инфраотряды Eotitanosuchida, Titanosuchida и Tapinocephalida. Горгонопсов М.Ф. Ивахненко разделил на инфраотряды Ictidorhinida, Gorgonopida и Estemmenosuchida.
По мнению Л.П. Татаринова (2009 г.), сближение эстемменозухов с какими-либо горгонопсами остается все же проблематичным. Разрастание наружного челюстного аддуктора на заглазничную кость — апоморфный признак, отсутствие которого должно характеризовать и прямых предков диноцелафов. Т. Кемп (1982 г.) поместил эстемменозухов в самое основание системы диноцефалов, ниже бритоподид (семейства Brithopodidae и Anteosauridae) и титанозухов (семейства Titanosuchidae и Tapinocephalidae), хотя Дж. Хопсон и Т. Баргхьюзен (1986 г.) считают возможной вторичную реверсию по этому признаку. Вместо эотерапсид Л.П. Татаринов (2009 г.) сохраняет более традиционное понятие эотериодонтов, но, в отличие от Б. Баттайла и М.В. Суркова (2000 г.), сокращает их объем, выделяя эту группу на основе фтинозухий и включая в нее под вопросом фтинозаврид.
Семейства биармозухид и эотитанозухид, которых многие исследователи (например, Кемп (1982 г.)) считают близкими к предкам горгонопсов, а М.Ф. Ивахненко в последних работах определяет как синонимы, описаны каждое по монотипическому роду из средней перми Пермской области; позднее со Среднего Урала были описаны еще 2-3 рода эотитанозухид. Собственно эотитанозухов М.Ф. Ивахненко относит к дейноцефалам отряда Titanosuchia, за исключением рода Kamagorgon, который он причисляет к фтинозухидам.
Для этой группы эотериодонтов характерно сочетание некоторых диноцефаловых и горгонопсовых признаков. В отличие от типичных териодонтов, восходящие пластины премаксилл у них необычно сильно вдаются между носовыми костями, чем биармозухи и эотитанозухи и напоминают диноцефалов. У биармозухид, как и у эотитанозухид, озубленные небно-птеригоидные бугры, характерные для горгонопсов, не развиты; некрупные зубы широко покрывают передние и, по-видимому, поперечные отростки птеригоидов, причем зубы на птеригоидных флангах никак не выделяются по размерам. Теменные кости очень короткие, и пинеальное отверстие, расположенное, как у горгонопсов, на костном бугре, относительно теменных костей выглядит очень крупным. По крайней мере у Biarmosuchus, в отличие от горгонопсов, наружный мыщелок квадратной кости расположен ниже внутреннего. Отмечается слабо выраженный передний сдвиг челюстного сустава. Наружная пластина угловой кости сдвинута к заднему концу челюсти сильнее, чем у горгонопсов. По строению посткрания, описанного у Biarmosuchus, отмечается близкое сходство с горгонопсами.
Трудности классификации фтинозухий отчетливо отражены в системе А. Бринка (1956 г.), который включил фтинозухий на правах подотряда в отряд горгонопсий. А. Бринк выделил в подотряде фтинозухий 7 семейств: Eotitanosuchidae, Biarmosuchidae, Phthinosuchidae, Phthinosauridae, Driveriidae, Mastersoniidae и Tappenosauridae. Биармозухиды включают 2 рода, а все остальные семейства — каждое только по 1 роду. По существу, А. Бринк относит к фтинозухиям всех не относящихся к диноцефалам примитивных хищных терапсид, которых считает отдаленными предшественyиками горгонопсов.
Л.П. Татаринов (2009 г.) считает фтинозухов весьма разнородной группой форм, филогенетически стоящих выше диноцефалов. В некоторых случаях намечаются их связи с териодонтами. Достаточно уверенно можно утверждать о связях рода Phthinosuchus с горгонопсами, которых целесообразно отделять от прочих териодонтов в качестве эотериодонтов. Фтинозухи могут включать и отдаленных предшественников высших териодонтов — терозухий. В качестве примера Л.П. Татаринов приводит фтинозавра, который может быть ближе к предкам тероцефалов или, что маловероятно, даже цинодонтов, чем к предкам настоящих горгонопсов.
Можно заключить, что среди эотериодонтов намечается дифференциация ряда групп более апоморфных терапсид, как это наблюдается и во многих других базальных группах рептилий.
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1 / Отв. ред. Ивахненко М. Ф. и Курочкин Е. Н. — М.: ГЕОС, 2008
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГеОС, 2009