Дромазавроиды — Dromasauroidea
Класс †Синапсиды (Synapsida)
Подкласс †Терапсиды (Therapsida)
Надотряд †Динообразные (Dinomorpha)
Отряд †Аномодонты (Anomodontia)
Подотряд †Хайнозавры (Chainosauria)
Надсемейство †Дромазавроиды (Dromasauroidea)
Дромазавроиды, или дромазавры (Dromasauroidea Broom, 1907), — парафилетическая группа, объединяющая наиболее базальных представителей подотряда хайнозавров (Chainosauria von Nopsca, 1923), условно рассматриваемая здесь в качестве надсемейства. Происходят из средней перми Африки. Установлены 1 семейство и 3 рода.

Реконструкция внешнего облика галепуса (Galepus jouberti). Автор — Mojcaj.
Размеры мелкие: общая длина составляет порядка зо см.
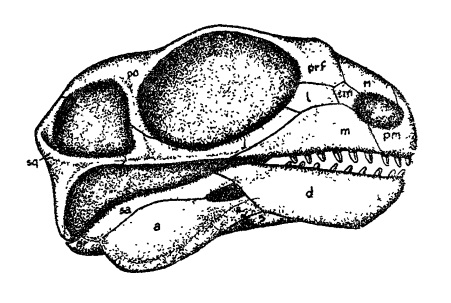
Прорисовка реконструированного черепа с нижней челюстью галепуса (Galepus jouberti).
Череп внешне напоминает таковой венюковиоидов. Он крупный относительно туловища, сравнительно короткий и довольно высокий. Лицевая часть укорочена и слегка наклонена вниз. Предчелюстная кость парная. Слезная кость большая, соприкасающаяся с септомаксиллой. Носовая кость контакт с септомаксиллой утратила. Глазница очень крупная. Заглазничная перегородка узкая. В отличие от венюковиоидов и дицинодонтов, заглазничная кость не тянется назад до чешуйчатой (по другим данным, эти кости контактируют).
Височное окно сравнительно короткое. Тонкая скуловая дуга поднята выше уровня нижнего края глазницы. Сохраняется задний отросток скуловой кости. Чешуйчатая кость узкая, ее нижний отросток резко удлинен. Как и у венюковиоидов, теменные кости короткие и широкие, большое пинеальное отверстие расположено на костном утолщении, непарная заднетеменная кость очень большая, а табличатые кости необычно протяженные. Затылок расширен умеренно. Челюстное сочленение вынесено далеко вниз.

Прорисовки реконструированных черепов с нижними челюстями дромазавров.
Неполное вторичное небо образовано предчелюстными костями. Подобно венюковиоидам, септомаксилла имеет длинный лицевой отросток. Поперечные отростки птеригоидов заметно редуцированы, направленные вперед и наружу. Небные зубы, как и у дицинодонтов, утрачены.
Нижнечелюстной симфиз слившийся. Нижняя челюсть без венечной кости, но венечный отросток, образованный зубной костью, иногда присутствует. Артикулярная кость с выпуклой сочленовной поверхностью, что свидетельствует о возможности совершать нижней челюстью пропалинальные движения. Аддукторная мускулатура распространяется из височного окна дорсально, а наружная поверхность нижней челюсти имеет борозду, указывающую на место прикрепления поверхностного слоя наружного аддуктора. В целом ключевые изменения челюстного аппарата, характерные для дицинодонтов, выражены мало.
Зубы колышковидные, отличаясь у разных родов по форме и количеству. Внутри группы наблюдается тенденция к постепенной утрате передних челюстных зубов. Клыкообразно увеличенных зубов не бывает. Роговой клюв также отсутствует.
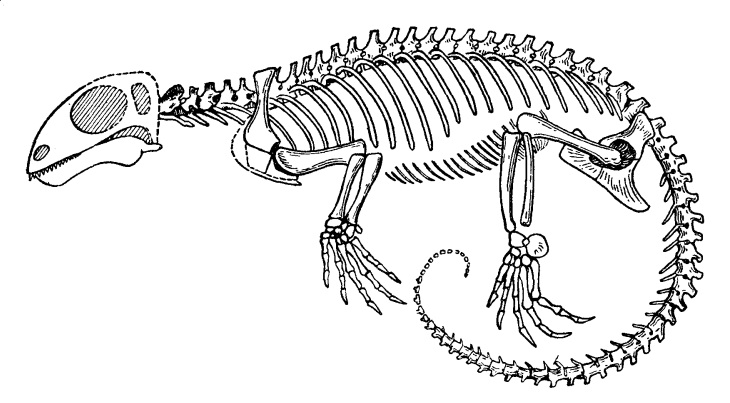
Старинная прорисовка реконструированного скелета галехира (Galechirus scholtzi).
Телосложение легкое, внешне напоминающее таковое ящериц. Шея хорошо обособлена и довольно длинная, из 5-7 позвонков. Туловище стройное. Спинных позвонков около 20. Поясничный отдел позвоночника не выражен. В крестце 2-3 позвонка. Хвост длинный, состоящий из 30-40 позвонков. Позвонки глубокоамфицельные, прободенные хордой. Шейные ребра короткие. Имеются многочисленные тонкие гастралии.
Лопатка узкая, без акромиона. Клейтрума нет. Коракоиды и межключица расширенные. Подвздошная кость очень слабо расширена. Седалищная кость направлена назад от уровня подвздошной. Запирательное отверстие лобковой кости увеличено, но тироидное отверстие отсутствует. Вентральный отдел тазового пояса пластинчатый. В отличие от дицинодонтов конечности длинные и тонкие. Плечевая кость с энтэпикондилярным отверстием. В кисти имеется 2 центральные косточки, в стопе — 1. Пяточная кость увеличена. Пальцы длинные, заканчивающиеся крупными когтевыми фалангами. Фаланговая формула, как и у дицинодонтов, маммального типа: 2 - 3 - 3 - 3 - 3.
Вероятно, дромазавры держались на достаточном отдалении от воды, например, в лесном подлеске. Судя по всему, они являлись достаточно подвижными. Питаться дромазавры могли насекомыми либо являлись всеядными, хотя растительноядность также не исключена.
Выказывалось предположение, что дромазавры представляют собой молодых особей каких-то более крупных аномодонтов, однако, поскольку взрослые особи известных видов не обнаружены, это предположение сомнительно.
Для надсемейства принимается следующий состав:
род †Галепусы (Galepus Broom, 1910);
род †Галехиры (Galechirus Broom, 1907);
семейство †Галеопсовые (Galeopsidae Broom, 1912);
род †Галеопсы (Galeops Broom, 1912).
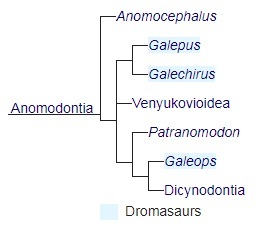
Сегодня принято рассматривать дромазавров как парафилетическую группу, родственную дицинодонтам. Ниже представлена обобщенная кладограмма, основанная на публикациях Модесто и Рубиджа (2000), Лю с соавт. (2009), Циснероса с соавт. (2011):
Ссылки
Дромазавры (Википедия) перейти
Dromasauroidea (Paleobiology Database) перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Иванов А. О., Черепанов Г. О. Ископаемые высшие позвоночные. — СПб.: Издательство Санкт-Петербургского университета, 2001
Ивахненко М. Ф. Тетраподы Восточно-Европейского плаката — позднепалеозойского территориально-природного комплекса. — Пермь, 2001
Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1 / Отв. ред. Ивахненко М. Ф. и Курочкин Е. Н. — М.: ГЕОС, 2008
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Ромер А. Ш. Палеонтология позвоночных. — М. — Л.: Государственное научно-техническое издательство нефтяной и горно-топливной литературы, 1939
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГеОС, 2009
Benton M. J., Shishkin M. A., Unwin D. M. The Age of Dinosaurs in Russia and Mongolia. — Cambridge University Press, 2000
Kemp T. S. The origin and evolution of mammals. — Oxford University Press, 2005