Двинии — Dvinia
Класс †Синапсиды (Synapsida)
Подкласс †Терапсиды (Therapsida)
Надотряд †Терозавры (Therosauria)
Отряд †Цинодонты (Cynodontia)
Семейство †Двиниевые (Dviniidae)
Род †Двинии (Dvinia)
Вид †Dvinia prima
Двинии (Dvinia Amalitzky, 1922) — монотипический род монотипического семейства двиниевых (Dviniidae Sushkin, 1928), происходящий из поздней перми Европы.

Реконструкция внешнего облика двинии (Dvinia prima). Автор — Р. Евсеев.
Синоним: Permocynodon Sushkin, 1927.
Единственный установленный вид — Dvinia prima Amalitzky, 1922 (тип). Его остатки обнаружены в местонахождении Соколки (Котласский район, Архангельская область, Россия). Они датируются поздней пермью (зона Scutosaurus, саларевская свита, вятский горизонт, верхневятский подъярус, татарский отдел). Синоним: Permocynodon sushkini Woodward, 1932.
Двиния была найдена В.П. Амалицким в 1899 г. в береговых отложениях реки Малая Северная Двина (отсюда название, данное первооткрывателем) у деревни Новинки под городом Котлас в костеносных линзах песков вместе с другими представителями позднепермской фауны.
Обнаружены неполная лицевая часть черепа крупной особи, почти полный череп небольшой особи, верхнечелюстная кость и задняя часть посткраниального скелета.


Череп с нижней челюстью двинии (Dvinia prima) из Соколковского местонахождения, Архангельская область, Россия.
Размеры некрупные: длина черепа достигает 12 см при максимальной ширине около 8,5 см, общая длина оценивается не более чем в 50 см. Несмотря на общую примитивность, это наиболее специализированный из позднепермских цинодонтов.
По своим очертаниям череп сильно напоминает таковой млекопитающего, но мозговая коробка очень мала. Морда сужена и сжата позади клыков. Глазницы крупные, обращены вперед и несколько наружу, отделены от височных окон узкой костной перемычкой. Задняя часть черепа сильно расширена за счет очень больших височных окон, длина которых составляет примерно половину общей длины черепа. Заднелобные кости отсутствуют. Сагиттальный гребень высокий и острый, начинается от задней части лобных костей и проходит по теменным. Пинеального отверстия нет. Затылок широкий, треугольный. Затылочных мыщелков 2.
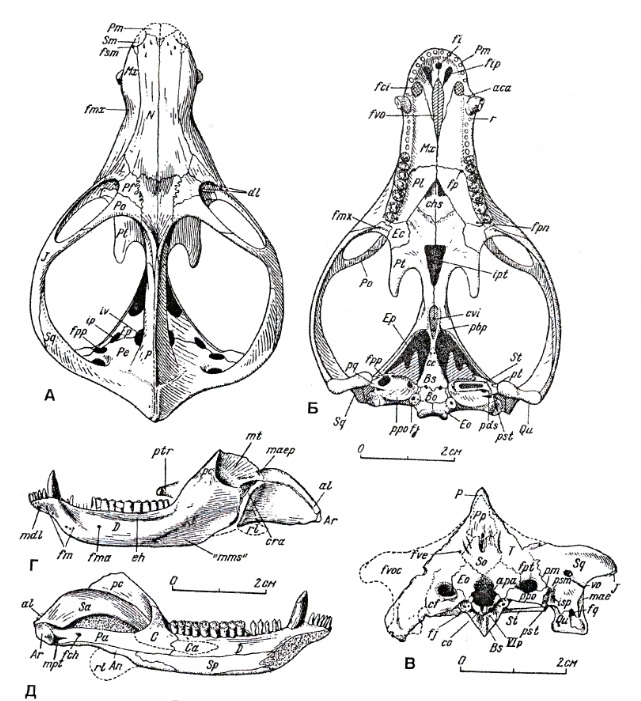
Прорисовки частично реконструированного краниального материала двинии (Dvinia prima) из Соколковского местонахождения, Архангельская область, Россия: А, Б, В — череп, Г, Д — нижняя челюсть.
В верхнечелюстной кости позади ноздри имеется глубокий желоб, куда при закрытой пасти входил клык нижней челюсти. Имеется отчетливо выраженное вторичное небо, образовавшееся за счет смыкания горизонтальных отростков небных и верхнечелюстных костей. Небо оканчивается на уровне VIII-IX-го заклыкового зуба, причем задние края образующих его небных костей расходятся от срединной плоскости под углом до 45 °. На уровне клыков во вторичном небе имеется узкая щель. Уникально сохранение неприкрытого почти парного сошника, глубоко погруженного между небными отростками премаксилл и максилл. Также сохраняется большая межптеригоидная яма. Поперечные отростки птеригоидов на конце сильно загнуты назад, что придает им сходство с птеригоидными отростками специализированных среднетриасовых хищных цинодонтов типа южноамериканского Chiniquodon. Эпиптеригоид соединяется с проотикой швом. Челюстное сочленение расположено немного ниже уровня затылочного.
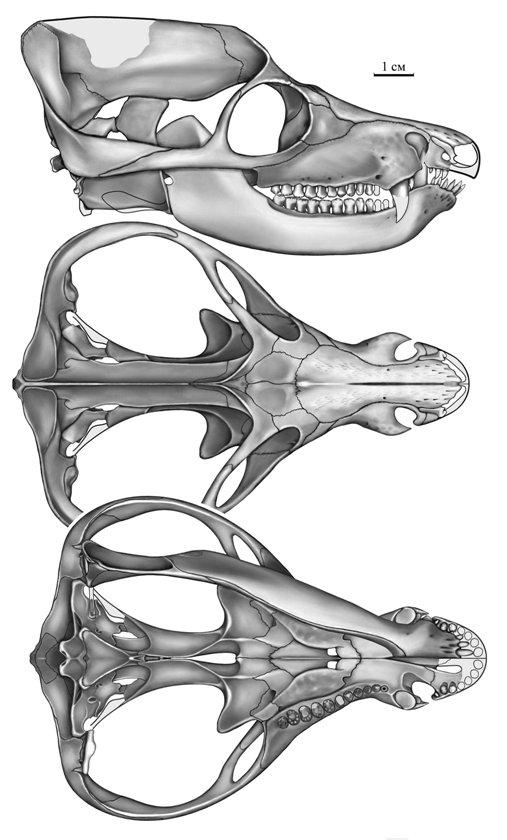
Прорисовки частично реконструированного черепа с нижней челюстью двинии (Dvinia prima) из Соколковского местонахождения, Архангельская область, Россия: сбоку, сверху и снизу.
Зубная кость сильно развита, с вытянутым вперед симфизным отделом и мощным венечным отростком. Необычной чертой для цинодонтов и вообще для терапсид является образование тонкого и удлиненного подбородочного выступа для резцовой части зубной кости, контрастирующего с массивным подбородком горгонопид. Помимо фасетки для m. temporalis, в задненижнем углу зубной кости появилась другая мышечная фасетка, возможно, для зачаточного m. masseter или для m. adductor mandibulae posterior. Задние кости нижней челюсти не редуцированы. Наружная пластина угловой кости пластинчатая, отогнута вниз и назад. Сочленовная кость занимает очень низкое положение на нижней челюсти, сравнимое с таковым у горгонопид и фтинозухид. На верхнезаднем крае сочленовной кости образовалось утолщение, которое при сомкнутых челюстях соприкасалось с утолщением на наружном крае чешуйчатой или квадратноскуловой костей, благодаря чему достигалось упрочнение сустава. Стремя мощнее, чем у обычных цинодонтов.
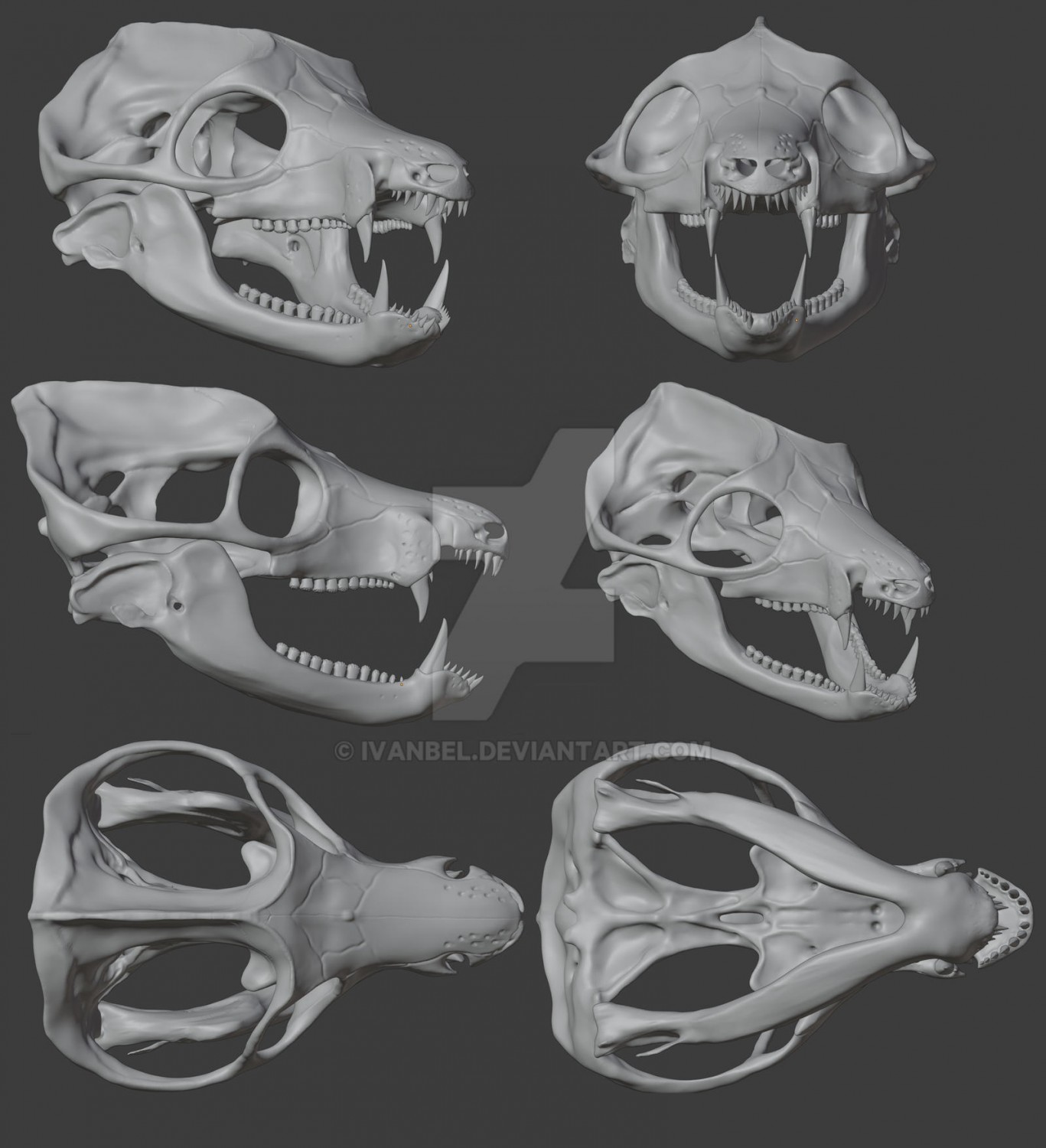
Компьютерные модели частично реконструированного черепа с нижней челюстью двинии (Dvinia prima) из Соколковского местонахождения, Архангельская область, Россия. Автор — И. Белоусов.
Зубная система очень своеобразная. В премаксилле 5 резцов, в нижней челюсти — 5-6. Клыков по 1 вверху и внизу. Заклыковых зубов в верхней челюсти 12, в нижней — 13-15. Резцы мелкие. Клыки большие, развиты намного сильнее, чем у Procynosuchus. Заклыковые зубы дифференцированы на простые, конической формы предкоренные (передние 6 зубов, из них 3 первых мелкие), а также резко уплощенные и поперечно расширенные многобугочатые коренные уникального типа. На верхних коренных главная вершинка большая и острая, заметно выступающая над уровнем придаточных, число которых не превышает 9-10. При этом наблюдается усложнение коронки и увеличение числа бугорков от передних зубов к задним, за исключением последнего. На нижних коренных главная вершинка почти не выступает над уровнем коронки, придаточные бугорки расположены 3 нечетко выраженными поперечными рядами, общее число бугорков достигает 20-22. На верхних коренных придаточные бугорки расположены по краям коронки вокруг главной вершинки, а на нижних — главным образом, внутрь от главной вершинки. Нижние заклыковые зубы расширены много больше верхних.
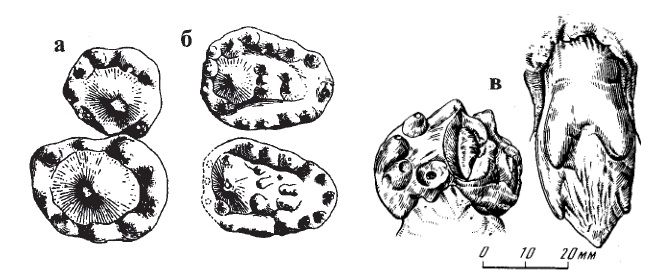
Прорисовки зубов двинии (Dvinia prima) из Соколковского местонахождения, Архангельская область, Россия: жевательные поверхности правых XII-XIII-х верхних и нижних зубов (а, б), задний заклыковой зуб сверху и снаружи (в).
В отличие от высших гомфодонтных цинодонтов и млекопитающих, при смыкании челюстей верхние и нижние ряды заклыковых зубов практически не окклюзировали. Обработка пищи производилась на поверхности небных отростков максилл и небных костей уплощенными нижнечелюстными зубами, а верхнечелюстные, имеющие высокую коническую вершинку, прижимали пищу к лабиальным площадкам нижних челюстей. Сильное развитие височных мышц и своеобразное строение квадратоартикулярного сустава, не допускавшее продольных или поперечных движений может указывать на то, что основной функцией заклыковых зубов являлось раздавливание достаточно твердой панцирной добычи, однако не вызывавшей сколько-нибудь значительного износа зубов. Последнее возможно при обработке очень тонкораковинных наземных моллюсков или панцирных насекомых.
Предкрестцовые позвонки амфицельные и несут ребра. Крестец включает 4 позвонка. Верхний отросток подвздошной кости сильно разрастается вперед. Обтураторное отверстие в тазе между лонной и седалищной костями таза очень большое (значительно крупнее, чем у Procynosuchus), почти круглое. Тазовая полость обширная. Общая форма вертлужной впадины сходна с таковой у млекопитающих. Бедренная кость стройная, с хорошо выраженной головкой полушаровидной формы, которая на 2/3 обращена своей сочленовной поверхностью медиально. В целом, морфологические особенности двинии указывают на достаточно высокий уровень ее специализации и значительную обособленность эволюционной линии от других цинодонтов.
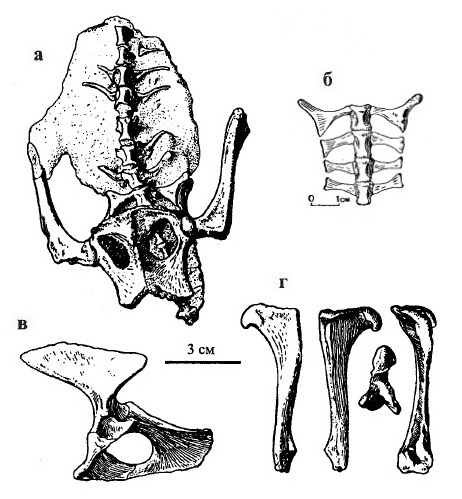
Прорисовки посткраниалного материала двинии (Dvinia prima) из Соколковского местонахождения, Архангельская область, Россия: крестцовая область снизу (а), крестец (б), левая половина таза сбоку (в), бедренная кость в различных видах (г).
Широкие скуловые дуги, очень крупные височные окна и сильное развитие венечного отростка нижней челюсти, вероятно, говорят о сильной и, возможно, хорошо дифференцированной челюстной мускулатуре. В отличие от некоторых тероцефалов, на лицевой части черепа отсутствуют отпечатки кровеносных сосудов, инфраорбитальный канал занимает более аборальное положение и предполагается наличие толстого слоя мягких тканей. Это подтверждается присутствием на верхнечелюстной кости области прикрепления мышцы поднимателя верхней губы (m. levator labii superior) с вероятными отпечатками губных мышц. Носовая полость разделена на этмоидную и максиллярную турбиналии, что наблюдается только у гомойотермных форм (птиц и млекопитающих) и связано с нагреванием вдыхаемого воздуха. Следовательно, можно полагать, что теплокровность была достигнута уже на уровне позднепермских цинодонтов.
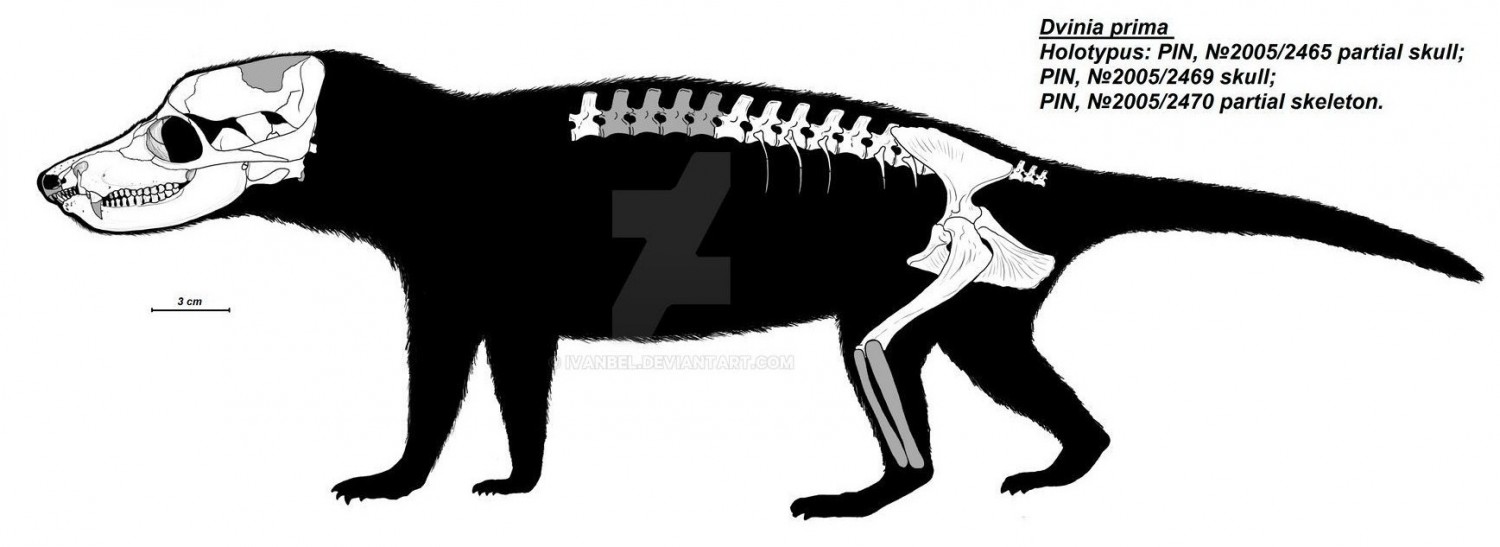
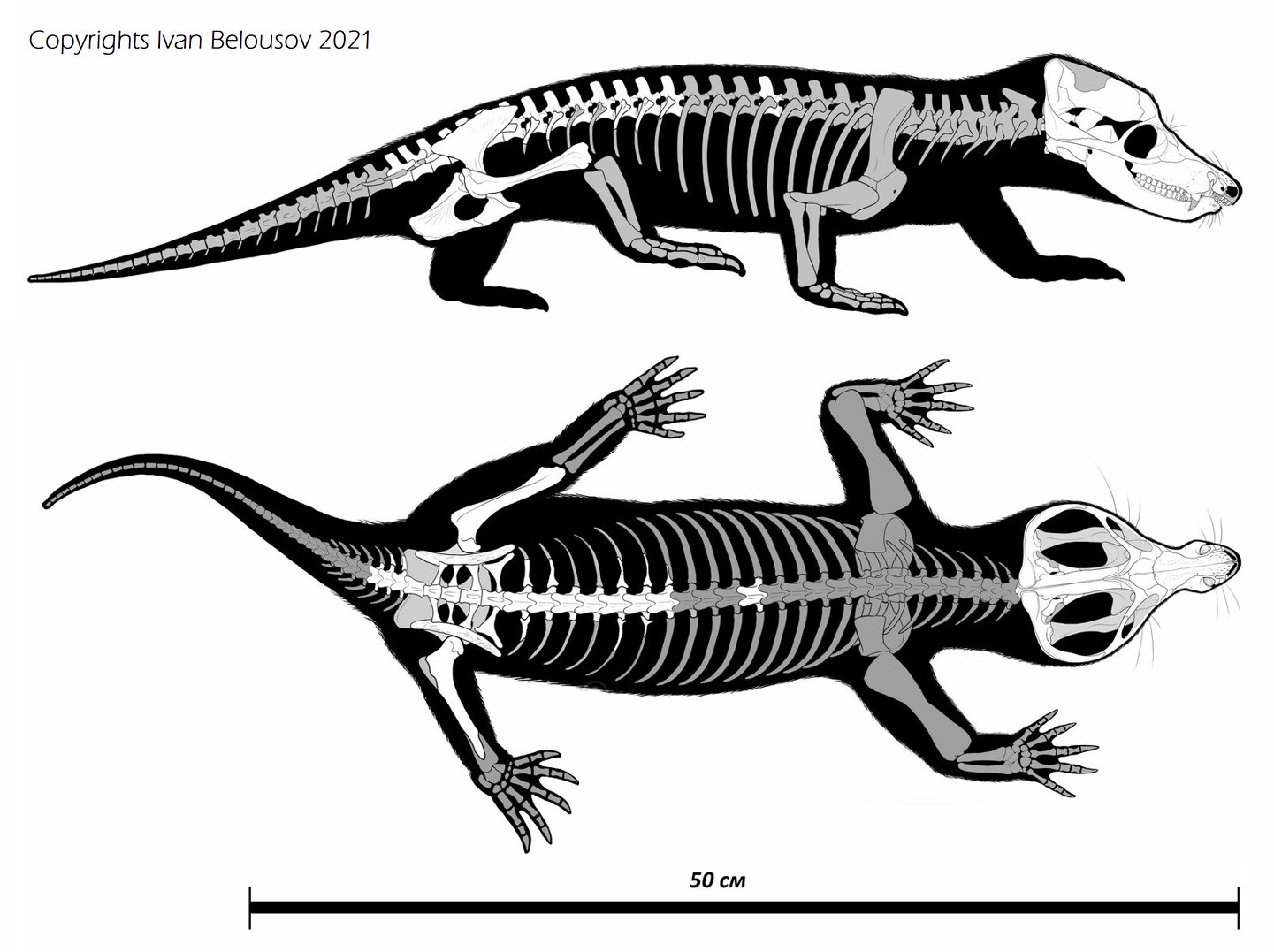
Прорисовки реконструированного частичного скелета двинии (Dvinia prima) из Соколковского местонахождения, Архангельская область, Россия. Автор — И. Белоусов.
Несмотря на то, что двиния по ряду признаков в строении черепа, таза и костей конечностей приближается к млекопитающим, наличие у нее нижней челюсти, состоящей из нескольких костей, а также отсутствие маммального прикуса говорят о примитивности этого цинодонта. С другой стороны, выраженная специализация заклыковых зубов двинии, которые не могут являться исходным состоянием для зубов млекопитающих, указывает на то, что она представляет собой боковую ветвь эволюции цинодонтов, не идущую в направлении к млекопитающим. Таким образом, при всем сходстве с млекопитающими, двиния также сильно от них отличается и не может являться для млекопитающих предковой формой.
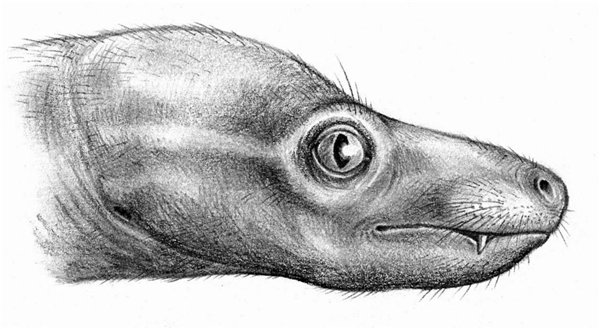
Портретная реконструкция двинии (Dvinia prima). Автор — А. Кац.
Данное животное входит в состав соколковского подкомплекса соколковского фаунистического комплекса наземных позвоночных. Его ископаемый материал встречается редко: в огромной коллекции из Соколков, насчитывающей порядка 2500 образцов разных форм, имеются остатки лишь 4 особей.
Обитая по соседству с многочисленными крупными хищниками этот небольшой цинодонт, скорее всего, вел довольно скрытный (возможно, сумеречный) образ жизни. Он занимал нишу наземного нанофага — насекомоядного, всеядного или даже полностью растительноядного.

Портретная реконструкция двинии (Dvinia prima). Автор — И. Белоусов.
Ссылки
Двиния (Википедия) перейти
Dvinia (Paleobiology Database) перейти
Gondwana Studios перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Буланов В. В. Северодвинские сокровища палеонтологии. — Архангельск: Территор. фонд информации по природ. ресурсам и охране окружающей среды МПР России по Арханг. Обл., 2007
Голубев В. К. Стратотипы и опорные разрезы верхней перми Поволжья и Прикамья. — Казань: Экоцентр, 1996
Ивахненко М. Ф. Тетраподы Восточно-Европейского плаката — позднепалеозойского территориально-природного комплекса. — Пермь, 2001
Ивахненко М. Ф. Эволюция позднепалеозойских тетрапод как эволюция их биоморф. // Эволюция биосферы и биоразнообразия / Ред. Рожнов С.В. — М.: Т-во научн. изд. КМК, 2006
Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1 / Отв. ред. Ивахненко М. Ф. и Курочкин Е. Н. — М.: ГЕОС, 2008
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Орлов Ю. А. В мире древних животных. — М.: Наука, 1989
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Очев В. Г. Еще не пришли динозавры. — Саратов: Научная книга, 2000
Павлова М. В. Палеозоология. Ч. 2. Позвоночные. — М.-Л.: Государственное издательство, 1929
Ромер А. Ш. Палеонтология позвоночных. — М. — Л.: Государственное научно-техническое издательство нефтяной и горно-топливной литературы, 1939
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГЕОС, 2009