Эндотиодоны — Endothiodon
Класс †Синапсиды (Synapsida)
Подкласс †Терапсиды (Therapsida)
Надотряд †Динообразные (Dinomorpha)
Отряд †Аномодонты (Anomodontia)
Подотряд †Хайнозавры (Chainosauria)
Инфраотряд †Дицинодонты (Dicynodontia)
Семейство †Эндотиодонтовые (Endothiodontidae)
Род †Эндотиодоны (Endothiodon)
Вид †Endothiodon bathystoma
Вид †Endothiodon tolani
Вид †Endothiodon mahalanobisi
Эндотиодоны (Endothiodon Owen, 1876) — род семейства эндотиодонтовых (Endothiodontidae Owen, 1876, non Lydekker, 1890), происходящий из средней - поздней перми Африки, поздней перми Азии и Южной Америки. Установлены 3 вида.
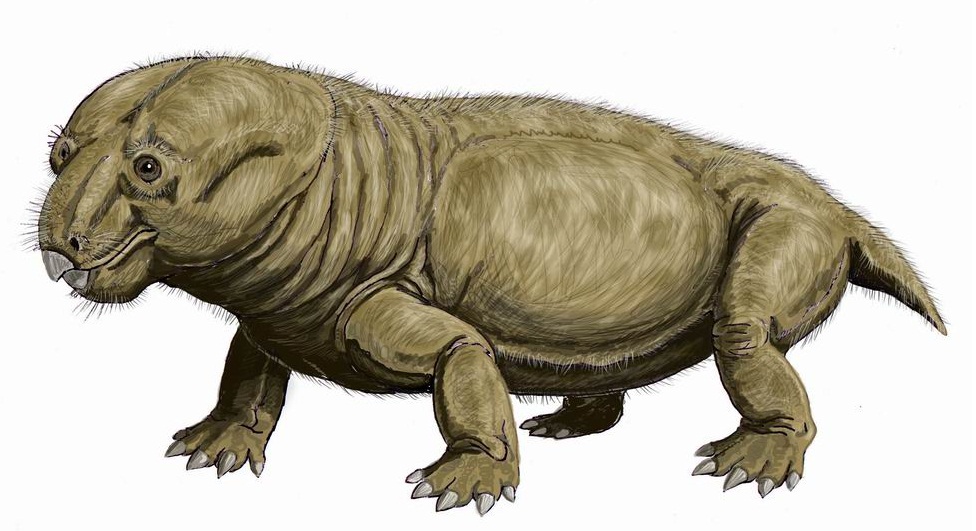
Реконструкция внешнего облика эндотиодона (Endothiodon bathystoma). Автор — Д. Богданов.
Синонимы: Esoterodon Seeley, 1895; Emydochampsa Broom, 1912; Endogomphodon Broom, 1932; Pachytegos Haughton, 1932.
Родовое наименование образовано греческими словами endothi (ἔνδοθῐ) — "внутри" и odon (ὀδών) — "зуб", по-видимому, намекая на расположение зубов внутри края верхнечелюстной кости.

Скелет эндотиодона (Endothiodon bathystoma) в блоке породы.

Череп с нижней челюстью эндотиодона (Endothiodon bathystoma).
Размеры от средних до крупных: у взрослых особей разных видов длина черепа варьирует, но не превышает 57 см, а общая длина составляет 1-2,5 м.
Череп относительно размеров тела очень большой. Он расширен сзади и почти треугольной формы, если смотреть сверху. Морда тупых очертаний с одним или тремя продольными гребнями на носовых костях. Небные кости, как правило, не соприкасаются с предчелюстными. Передняя часть нижней челюсти изогнута вверх, образуя как бы перевернутый клюв. Предчелюстные кости и небо формируют свод, куда помещается заостренный конец нижней челюсти.
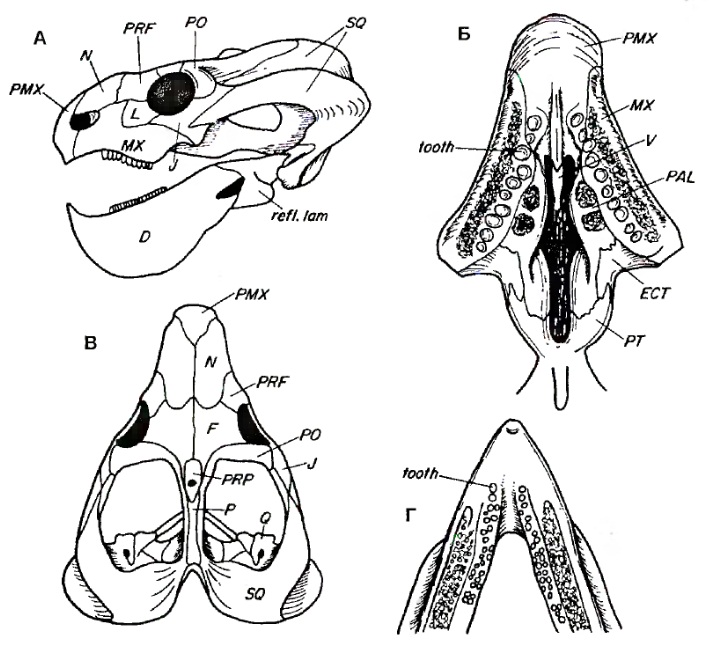
Прорисовки реконструированного черепа с рнижней челюстью эндотиодона (Endothiodon bathystoma): А — сбоку, Б — сверху, В — снизу, Г — нижняя челюсть сверху.
Клыки в большинстве случаев утрачены. В задней части верхней челюсти и средней части нижней находятся многочисленные мелкие зубы — до 11 вверху и до 28 внизу. Зубы длинные и тонкие, отличаясь в верхней и нижней челюстях как морфологически, так и по способу замены. Верхние зубы взрослых особей, как правило, крупнее (5-9 мм), чем зубы нижней челюсти (менее 5 мм), и имеют зазубрины на переднем крае, тогда как нижние зазубрены по заднему краю. Дистальная часть каждого нижнего зуба сжата с боков и имеет несколько грушевидную форму в поперечном сечении.

Намного более грацильная нижняя челюсть молодых особей содержит один функциональный ряд из 5-6 зубов. Зубы маленькие, конические и заостренные. Дистальная поверхность покрыта зачаточными зубчиками. Ювенильные зубы устроены гораздо проще и больше напоминают зубы плотоядного животного, чем фитофага. Возможно, различное строение зубов обусловлено изменением способа питания от насекомоядного или всеядного в молодом возрасте до растительноядного во взрослом. Зубная морфология менялась по мере увеличения размеров животного, все более приспосабливаясь к растительному рациону.
Верхние зубы расположены внутри края верхнечелюстной кости, образуя один неровный ряд. Как правило, также имеются неправильные скопления зубов на небных отростках максилл. Нижние зубы располагаются на зубной кости тремя диагональными рядами. В каждом ряду самый передний (крайний) зуб является наиболее старым, а задний — самым молодым. На протяжении жизни шло активное замещение зубных рядов. Зубы верхней челюсти не противопоставлены зубам нижней.
Верхнечелюстной фрагмент эндотиодона (Endothiodon bathystoma), виден неправильный зубной ряд.
На небе имеется два четко ограниченных участка, пронизанных мелкими отверстиями. Эти участки, вероятно, имели роговое покрытие. Широкая борозда, проходящая по зубной кости вдоль наружной стороны зубного ряда, по-видимому, также была покрыта роговым слоем. Судя по всему, верхние зубы входили в бороздку рогового клюва, лежавшую снаружи от нижних зубов. Наружный ряд нижних зубов противопоставлялся роговому покрытию неба.
Туловище массивное, бочкообразное. Укороченные конечности мощные, расположенные по бокам тела. Хвост очень короткий.
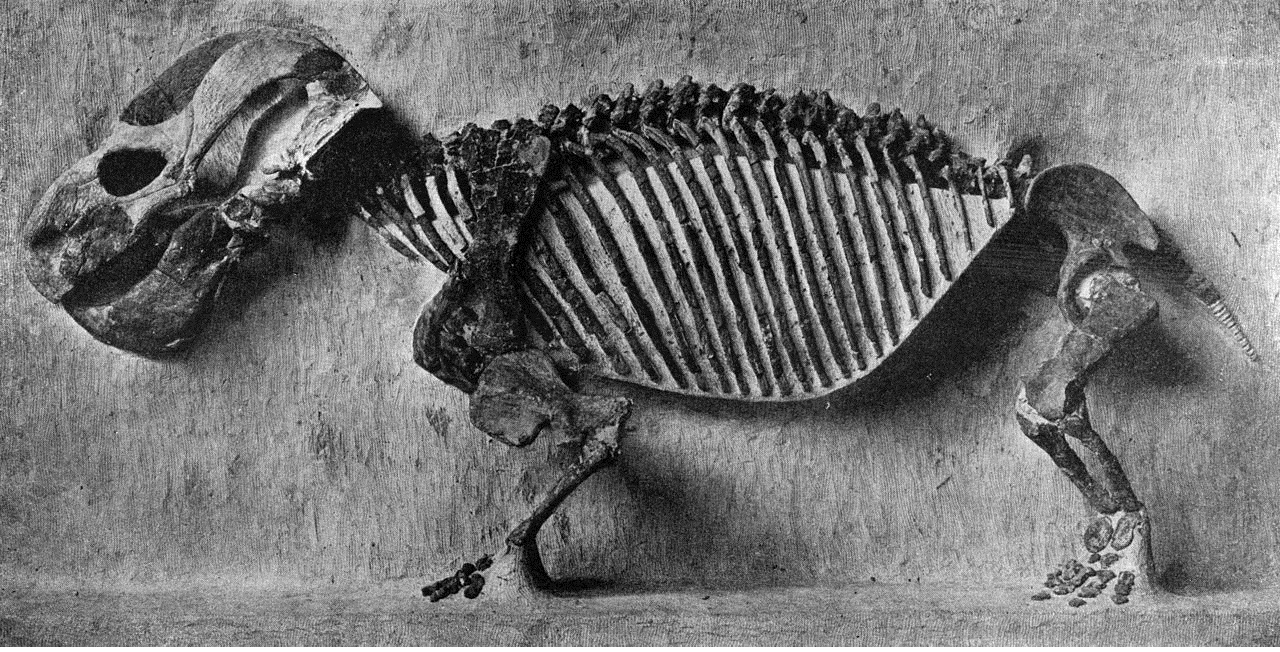


Скелет эндотиодона (Endothiodon bathystoma).
Endothiodon bathystoma Owen, 1876 (тип) — типовой вид, известный из поздней перми (зоны Tropidostoma - Cistecephalus, вучапинский ярус, лопинский отдел) ЮАР (Северная Капская, Западная Капская и Восточная Капская провинции) и поздней перми (примерное соответствие зоне Cistecephalus, вучапинский ярус, лопинский отдел) Танзании (область Рувума). Форма, обозначенная Endothiodon cf. bathystoma, происходит поздней перми (примерное соответствие зоне Cistecephalus, вучапинский ярус, лопинский отдел) Малави (Северная провинция).
Синонимы: Endothiodon uniseries Owen, 1879; Endothiodon platyceps Broom, 1912; Emydochampsa platyceps (Broom, 1912); Endothiodon whaitsi Broom, 1912; Esoterodon whaitsi (Broom, 1912); Endothiodon paucidens Broom, 1915; Esoterodon paucidens (Broom, 1915); Endothiodon seeleyi Broom, 1915; Endothiodon angusticeps Broom, 1915; Esoterodon angusticeps (Broom, 1915); Endogomphodon minor Broom, 1932; Endothiodon minor (Broom, 1932); Endothiodon crassus Broom, 1932; Endogomphodon crassus (Broom, 1932); Pachytegos stockleyi Haughton, 1932; Emydochampsa oweni Broili et Schröder, 1936; Endothiodon oweni (Broili et Schröder, 1936); Endogomphodon oweni (Broili et Schröder, 1936).
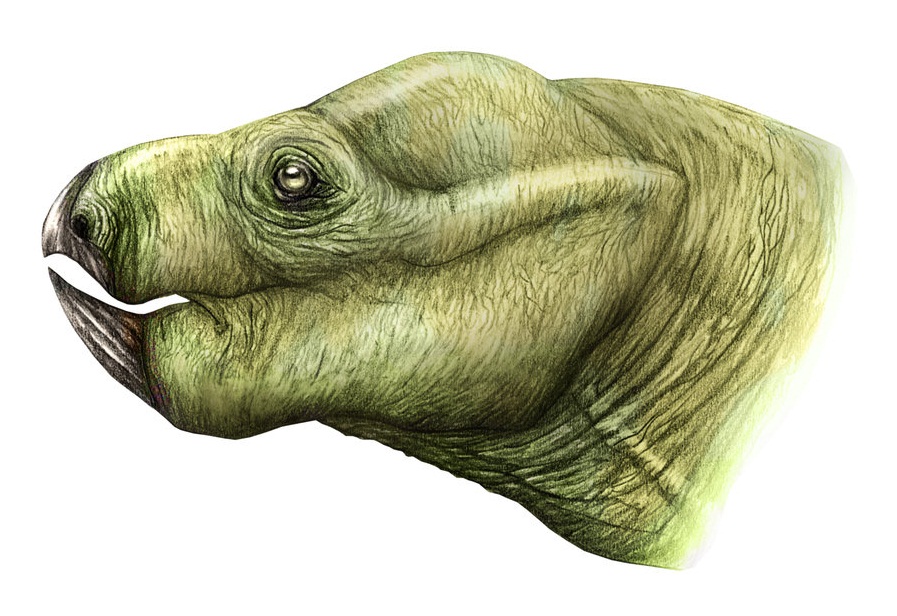
Портретная реконструкция эндотиодона (Endothiodon bathystoma).
Тип рода был описан Р. Оуэном в 1876 г. по остаткам, обнаруженным в позднепермских отложениях южноафриканской группы Бофорт, входящей в формацию Карру. Материалом для первоописания стала передняя часть морды и соответствующий ей нижнечелюстной фрагмент. Впоследствии здесь было собрано большое количество образцов, в частотности в 1915 г. Р. Брум обнаружил первый частичный скелет. Вслед за типовым из Южной Африки были описаны многочисленные новые виды и рода, объединяемые в подсемейство Endothiodontinae.
В 1964 г. Б. Кокс разобрал все таксоны эндотиодонтин и выявил несостоятельность диагностических признаков, разделявших рода Endothiodon, Esoterodon, Emydochampsa, Endogomphodon и Pachytegos. Все они были сведены в род Endothiodon. Располагая первоначально более чем полутора десятком видов эндотиодонтин, Б. Кокс смог сократить их число до трех — Endothiodon uniseries, E. bathystoma и E. whaitsi. Однако поскольку различия между этими видами заключаются главным образом в общих размерах, можно с высокой долей уверенности предположить, что они представлять собой единый онтогенетический ряд типового вида, а не три отдельных таксона.
На носовых костях имеются три продольных гребня. Пинеальное отверстие круглой формы. Клыки полностью отсутствуют.
Endothiodon tolani Cox et Angielczyk, 2015 — вид, известный из средней перми (кептенский ярус (?), гваделупский отдел) Танзании (область Рувума). Остатки данного таксона были обнаружены еще в 1963 г. От остальных видов его отличает отсутствие шишковидного выступа и сохранение пары верхних клыков сбоку от зубного ряда.
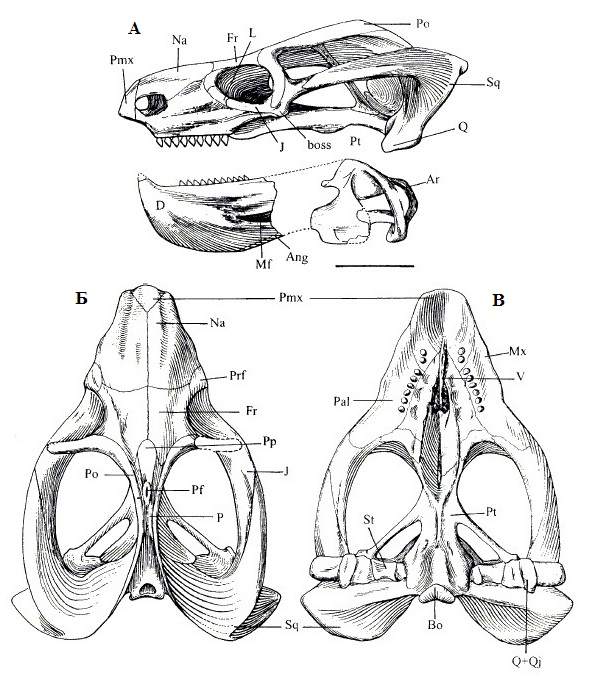
Прорисовки реконструированного черепа с нижней челюстью эндотиодона (Endothiodon mahalanobisi): А — сбоку, Б — сверху, В — снизу.
Endothiodon mahalanobisi Ray, 2000 — вид, известный из поздней перми (примерное соответствие зонам Tropidostoma / Cistecephalus, вучапинский ярус, лопинский отдел) Индии (штат Андхра-Прадеш). Найдены остатки как взрослых, так и молодых особей. Размеры небольшие. Носовые кости несут единственный продольный гребень (по сравнению с тремя у других видов). Шишковидный выступ расположен относительно низко. Предлобная кость вздута. Удлиненное пинеальное отверстие находится на невысоком вздутии, располагаясь посередине межвисочных гребней. Нижнечелюстной симфиз тонкий.
Endothiodon sp. — серия находок, происходящая из поздней перми (зона Cistecephalus, вучапинский ярус, лопинский отдел) Замбии (Южная провинция).
Endothiodon sp. — форма, известная из средней или поздней перми (формация Рио-ду-Расто, кептенский / вучапинский ярус, гваделупский / лопинский отдел) Бразилии (штат Парана). В 1970-х гг. здесь были найдены череп и нижняя челюсть, которые отнесли к роду Endothiodon. Это был первый дицинодонт пермского возраста, обнаруженный в Южной Америке. В 2013 г. А. Бус заново изучил образцы и подтвердил их принадлежность данному роду.

Группа эндотиодонов (Endothiodon bathystoma). Автор — Г. Угуэто.
Как показывает большое количество и обширная география находок остатков эндотиодонов, они были весьма обильны на значительной части Гондваны. По преобладанию остатков этого рода дицинодонтов средняя зона позднепермских отложений в Южной Африке, следующая за зоной Tapinocephalus, была названа зоной Endothiodon. Однако, поскольку их остатки не ограничивались одноименной зоной, ее переименовали в зону Tropidostoma (по названию другого дицинодонта).

Кормящиеся эндотиодоны (Endothiodon bathystoma).
Первоначально предполагалось, что эндотиодоны выкапывали из почвы корни с помощью своего клюва. В дальнейшем эта гипотеза была признана неправдоподобной, поскольку наружные ноздри на морде этих животных направлены вперед. Сегодня полагают, что эндотиодоны кормились сочными растениями, захватывая их клювом и перетирая большой специализированной пастью. Тафономическая реконструкция местообитаний подтверждает наличие здесь в поздней перми густой прибрежной растительности. Также существует предположение о питании эндотиодонов шишками хвойных.
Лиценопс (Lycaenops sp.) пытается найти угол атаки на эндотиодона (Endothiodon sp.). Автор — В. Синкконен.
Ссылки
Эндотиодон (Википедия) перейти
Endothiodon (Paleobiology Database) перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГеОС, 2009
Kemp T. S. The Origin and Evolution of Mammals. — New York: Oxford University Press, 2005