Ехидновые — Tachyglossidae
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Йинотерии (Yinotheria)
Клада Австралосфениды (Australosphenida)
Надотряд Прототерии (Prototheria)
Отряд Однопроходные (Monotremata)
Надсемейство Орниторинхоиды (Ornithorhynchoidea)
Семейство Ехидновые (Tachyglossidae)
Ехидновые (Tachyglossidae Gill, 1872) — семейство надсемейства орниторинхоидов (Ornithorhynchoidea Flannery, McCurry, Rich, Vickers-Rich, Smith et Helgen, 2024), происходящее из плиоцена - современности Австралии и позднего плейстоцена - современности Новой Гвинеи. Установлены 1-2 ископаемых и 2 современных рода.
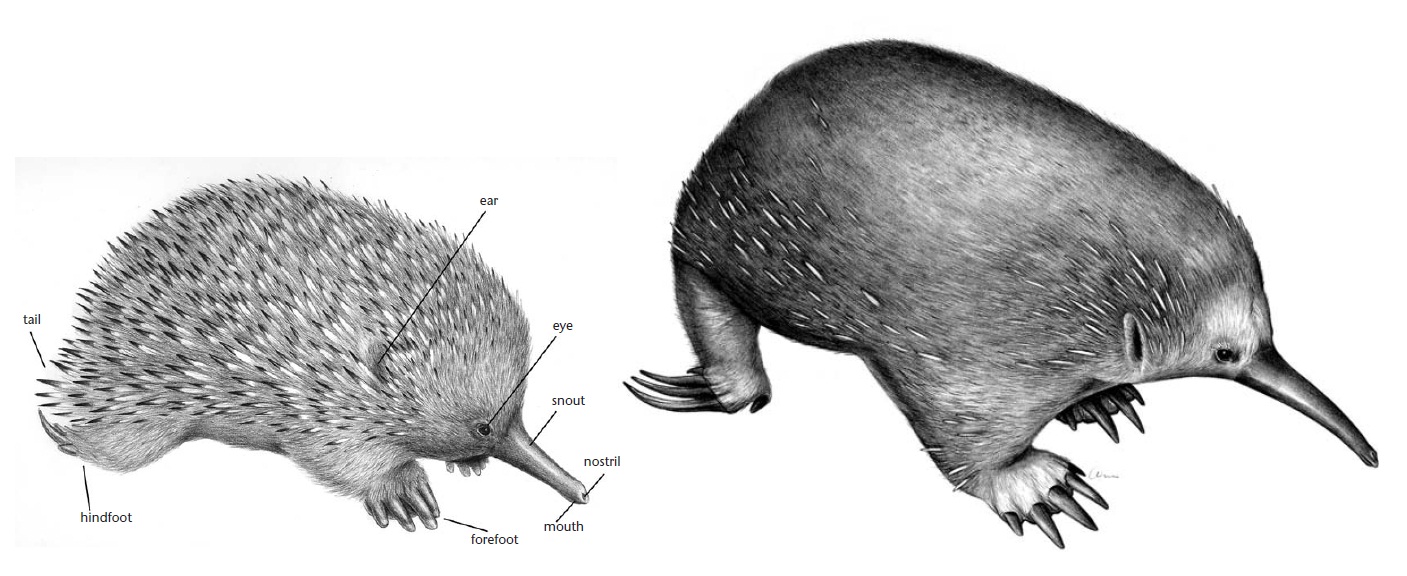
Сравнение внешнего облика австралийской ехидны (Tachyglossus aculeatus) и проехидны Бартона (Zaglossus bartoni).
По сравнению с утконосовыми (Ornithorhynchidae), ехидновые более специализированы и сохранили меньше сходных черт с общими предками. Длина тела современных представителей этой группы варьирует в пределах 30-76 см. Самцы обычно крупнее самок.
Относительно маленькая, продолговато-округлая голова резко отграничена от вытянутого в длину, цилиндрического рыльца-рострума. Оно выпуклое сверху, уплощенное снизу, довольно широкое в основании, к концу сужается и заканчивается тупо. На самом конце рыльца — там, где покрывающая его голая кожа становится мягче — расположены сравнительно большие овальные ноздри и очень маленькая и узкая ротовая щель. Рыльце является приспособлением для поиска добычи в щелях и лесной подстилке и по своей форме отличается у представителей разных родов. Маленькие выпуклые глаза лежат по бокам головы. Они прикрываются толстыми морщинистыми веками и, в отличие от утконоса, лишены мигательных перепонок. Наружные слуховые проходы представляют собой широкие хрящевые воронки, окруженные мышцами. Большие ушные отверстия имеют форму вертикальных щелей, скрытых в шерсти и иглах. Ушные раковины отсутствуют или очень невелики.
Шея короткая, снаружи практически незаметная. Туловище компактное, приземистое, несколько уплощенное, с куполообразной спиной и плоским брюхом. Мощные конечности относительно короткие (австралийская ехидна) или довольно высокие (проехидны), обе их пары имеют примерно одинаковую длину. Передние конечности прямые, задние отодвинуты назад и вывернуты наружу. Широкие лапы несут свободные, но не вполне подвижные пальцы числом от 3 до 5. Они вооружены длинными, широкими и крепкими когтями, особенно развитыми на передних лапах. Эти когтистые лапы хорошо приспособлены для рытья. II-й и III-й пальцы задних лап несут очень длинные изогнутые когти, которые благодаря особенностям строения тазового пояса и задних конечностей могут использоваться для чистки меха и игл практически на любой части тела. Хвост сильно укороченный, прямой и притупленный на конце.
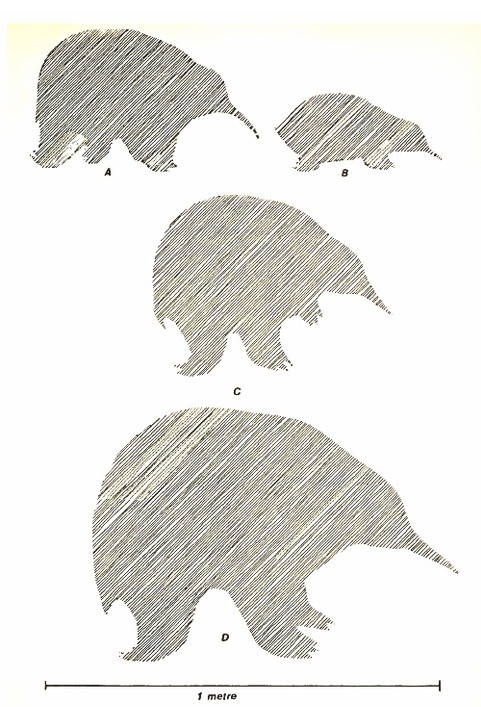
Относительные размеры ехидн: A — проехидна Брюйна (Zaglossus bruijni); B — австралийская ехидна (Tachyglossus aculeatus); C — мегалибвилия массивная (Megalibgwilia robusta) и мегалибвилия Рамсая (Megalibgwilia ramsayi); D — муррейглосс (Murrayglossus hacketti).
Кожа имеет значительную толщину. В ее среднем и глубинном слое проходят пучки поперечнополосатых мышц. Гладкие мышцы в коже отсутствуют. Потовые железы на теле имеются (проехидны) или отсутствуют (австралийская ехидна). Сальные железы располагаются только у волосяных сумок. В отличие от утконоса, задняя часть головы, спина, бока и обычно хвост сверху покрыты иглами — преобразованными волосками. Длина игл достигает 6 см, в поперечнике они округлые, снаружи гладкие, с толстым корковым слоем и полые внутри. Между иглами растет шерсть — такая же, как на большей части головы, дистальных отделах конечностей и брюхе, где игл нет. Шерсть может быть длиннее игл. Волосы грубые, без видимого сердцевинного слоя. Иглы и волосы окрашены в черные, бурые и желтоватые тона. Поднятие иголок осуществляется за счет сокращения толстого слоя подкожной мускулатуры, в котором они закреплены.
По наличию на внутренней стороне пяток задних лап остроконечных роговых шпор взрослые самцы легко отличаются от самок. Шпора полая и соединена каналом со слабо развитой железой, расположенной в области подколенной ямки. В отличие от утконоса, в качестве ядовитого аппарата шпора и железа не действуют. Ее секрет используется для других целей — предположительно, для запаховой коммуникации, либо выполняет какую-то еще неизвестную функцию. Зачатки шпор имеются у детенышей обоих полов, но у самок они почти всегда исчезают до наступления половой зрелости, а их железы так и остаются нефункциональными. Иногда шпоры отсутствуют и у взрослых самцов.
Носовые, септомаксиллярные, предчелюстные и верхнечелюстные кости образуют длинное и тонкое рыльце. У его переднего конца лежит округлое отверстие наружных ноздрей, всецело ограниченное септомаксиллами. Отверстия на морде для прохождения ответвлений тройничного нерва более многочисленны, чем у утконосов, но гораздо меньшие по размерам. Края верхнечелюстных костей узкие. Зубы совершенно отсутствуют, они не обнаружены даже на эмбриональных стадиях развития. Нет также зубной пластинки. Костное небо еще длиннее, чем у утконосов. Массивный эктоптеригоид, слитый с задним отделом небной кости, простирается далеко назад и вместе с продолжением каменистой кости формирует часть стенки барабанной полости, однако сам слуховой барабан не образуется. Барабанная кость крюковидно изогнута. Молоточек и наковальня соединяются друг с другом анкилозом и с помощью дополнительного отростка плотно прикреплены к каменистому отделу височной кости черепа. Барабанная кость плотно присоединяется, а часто даже срастается с отростком молоточка. Скуловые дуги очень тонкие. Сочленовная ямка для нижней челюсти мелкая, отросток за ней отсутствует.
Нижняя челюсть сильно редуцирована и является самой слабой среди млекопитающих. Она немного короче верхней челюсти и состоит из 2 длинных, тонких, сужающихся к концу и слегка изогнутых стержней, свободно соединенных в симфизе. Суставный и угловой отростки выражены плохо, а венечный отросток почти исчез. Массетерная ямка полностью утрачена. С верхней челюстью нижнюю соединяют сильные продольные краевые связки, простирающиеся почти до самого кончика морды. Вследствие этого нижнечелюстные ветви не движутся в верхне-нижнем направлении, а вращаются вокруг своих продольных осей — уникальный механизм открытия рта, известный только у ехидновых. Посткраниальный скелет гораздо массивнее, чем у утконосовых. Характерна особенно мощная мускулатура.
Язык тонкий, червеобразный. У ехидны он может далеко высовываться из ротового отверстия и изгибаться в разные стороны. У проехидн язык высовывается изо рта всего на несколько сантиметров, хотя сам по себе он длиннее, чем у ехидны. На передней части языка имеются роговые шипики, размещение которых неодинаков у обоих родов в связи с различной пищевой специализацией. Сзади в верхней части языка расположена подушечка из мелких ороговевших шипов. Пища измельчается благодаря тому, что они трутся о противостоящие роговые шипы на небных складках. Слюнные железы очень большие, особенно подъязычные. Они выделяют клейкую слюну, обильно покрывающую язык. У грудины имеется мечевидный отросток, к которому прикреплены сильно развитые язычные мышцы.
Мозг значительно крупнее мозга утконоса, со значительным развитием обонятельных долей (макросматический). Большие полушария округлые, со значительным количеством извилин, особенно у проехидн. Зрение слабое, но слух и обоняние развиты хорошо. В коже на дистальной части рыльца залегают рецепторы электрической чувствительности, но в гораздо меньшем числе, чем на клюве у утконоса. Полагают, что они представляют собой специализированные эккриновые железы. Электрорецепторы помогают обнаруживать слабые электрические поля, производимые почвенными беспозвоночными, но важной роли в жизнедеятельности, по-видимому, не играют. Кроме того, в коже рыльца присутствуют механические рецепторы и несэнсорные эккриновые железы, вырабатывающие серозно-слизистую смазку.

Самец австралийской ехидны (Tachyglossus aculeatus) с эрегированным пенисом.
Функционируют оба яичника. Сумка у самок едва заметна большую часть года. Перед началом сезона размножения складки кожи и подкожные мышцы на обеим сторонах живота увеличиваются, образуя незамкнутую сумку полулунной формы. Ее отверстие открывается вперед, а внутри по бокам находятся 2 млечных поля — небольшие углубления, покрытые волосками. Во время вынашивания яйца за счет сильного прилива крови температура сумки становится выше, чем в остальном теле. По мере роста детеныша сумка расширяется, а по выходе его вновь сглаживается. У самца имеется длинный пенис, головка которого разделяется на 4 симметричных лопасти. В кариотипе обоих современных родов 63 (самец) или 64 (самка) хромосомы.
Обитатели лесных, парковых и кустарниково-степных ландшафтов, в горы поднимаются до 4000 м над ур. моря. Наземные, медлительные животные. Имеют своеобразную походку вразвалочку. В отличие от касающегося земли брюха утконоса, в данном случае оно приподнято достаточно высоко. Активны преимущественно в сумеречное и ночное время суток. День проводят в убежищах — пустотах между камнями и корнями деревьев, полых бревнах или в норах, которые выкапывают сами. Могут преодолевать различные препятствия — завалы из древесных стволов, нагромождения камней и др. В случае необходимости переплывают довольно большие водоемы. Австралийская ехидна при опасности быстро зарывается в землю или сворачивается в колючий шар.

Ориентация и форма рыльца ехидн, отражающие их пищевую специализацию: а — мегалибвилия массивная (Megalibgwilia robusta); б — проехидна Брюйна (Zaglossus bruijni); в — австралийская ехидна (Tachyglossus aculeatus).
Питаются взрослыми насекомыми и их личинками, многоножками, червями, моллюсками и другими почвенными беспозвоночными. Корм извлекают из щелей и лесной подстилки длинным и липким языком. Для представителей разных родов характерна различная пищевая специализация и предпочитаемые местообитания. Обмен веществ невысокий. В любое время года при недостатке кормов способны впадать в спячку, которая сопровождается снижением температуры тела и интенсивности метаболизма. Ведут одиночный образ жизни; территориальность выражена слабо. Социальные взаимодействия проще, чем у большинства других млекопитающих. Максимальная зарегистрированная продолжительность жизни — свыше 50 лет.
Ареал охватывает Австралию с Тасманией и Новую Гвинею.
Состав семейства следующий:
род Ехидны (Tachyglossus Illiger, 1811);
род Проехидны (Zaglossus Gill, 1877);
? род †Мегалибвилии (Megalibgwilia Griffiths, Wells et Barrie, 1991);
род †Муррейглоссы (Murrayglossus Flannery, Rich, Vickers-Rich, Ziegler, Veatch et Helgen, 2022).
В отличие от утконосов, ехидны появляются в палеонтологической летописи очень поздно — только в плиоцене. У них отсутствуют зубы, которые в большинстве случаев лучше всего сохраняются в ископаемом состоянии. На основе морфологической консервативности и высокой специализации долгое время считалось, что эволюционные линии ехидн и утконосов разошлись не позднее среднего мела. Однако относительно свежие молекулярно-генетические исследования свидетельствуют, что это разделение произошло гораздо позже — ориентировочно, в палеогене. Следуя этой точке зрения, раннемеловые однопроходные оказываются скорее не примитивными родственниками современного утконоса, а базальными формами, в равной степени отдаленными как от ехидн, так и от настоящих утконосов. Таким образом, общий предок ехидны и утконоса мог в значительной мере напоминать последнего и много времени проводить в воде.
Вполне вероятно, что в дальнейшем ехидны вторично перешли к наземному образа жизни. На наличие у них полуводных предков могут указывать обтекаемые контуры тела, не препятствующие передвижению в воде, разведенные в стороны ноги с большими плоскими лапами, которые в прошлом могли служить для плавания, усиленное вращение плеча вокруг его продольной оси, что могло обеспечивать эффективные гребные движения передних лап, вывернутые назад задние конечности, которые некогда могли выполнять функцию рулей, сходство рыльца на определенном этапе эмбрионального развития с утконосим клювом, а также присутствие на нем электрорецепторов, которые могли использоваться для поиска пищи в водной среде. Впрочем, ехидны и сегодня хорошо плавают, если у них возникает такая потребность. Возвращение на сушу наверняка должно было привести к конкуренции с сумчатыми, но ехиднам все-таки удалось прочно закрепиться в невостребованной до них нише мирмекофагов.
В ходе эволюции ехидны независимо обрели многие морфо-физиологические черты, общие с другими мирмекофагами — южноамериканскими муравьедами (Vermilingua) из числа плацентарных. У представителей обеих групп передняя часть черепа вытянулась в трубкообразное, лишенное зубов рыло. Параллельно данному процессу происходила редукция нижней челюсти. Развился также длинный червеобразный язык, крупные слюнные железы, вырабатывающие липкую слюну, и мощные когти передних лап для раскапывания либо проламывания стенок гнезд колониальных насекомых.
Состав
Echidna (Wikipedia) перейти
Tachyglossidae (Animal Diversity Web) перейти
Tachyglossidae (Paleobiology Database) перейти
Литература
Grzimek's animal life encyclopedia. Volumes 12-16 / Ed. by Hutchins M., Kleiman D. G., Geist V., McDade M. C. — 2rd ed. — Farmington Hills, MI: Gale Group, 2003
Mammal Species of the World. A Taxonomic and Geographic Reference / Ed. by Wilson D. E., Reeder D. M. — 3rd ed. — Baltimore: Johns Hopkins University Press, 2005
Musser A. M. Furry Egg-layers: Monotreme Relationships and Radiations. In Merrick J. R., Archer M., Hickey G. M., Lee M. S. Y. (eds). Evolution and Biogeography of Australasian Vertebrates. — Sydney: Australian Scientific Publishing, 2006
Nowak R. M. Walker’s Mammals of the World. Monotremes, Marsupials, Afrotherians, Xenarthrans, and Sundatherians. — Baltimore: Johns Hopkins University Press, 2018
Nowak R. M., Paradiso J. L. Walker’s Mammals of the World. — 4nd ed. — Baltimore/London: Johns Hopkins University Press, 1983
Rose K. D. The beginning of the age of mammals. — Baltimore: The Johns Hopkins University Press, 2006