Гапалодекты — Hapalodectes
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Клада Скротиферы (Scrotifera)
Клада Феревунгуляты (Fereuungulata)
Клада Эвунгуляты (Euungulata)
Клада Параксонии (Paraxonia)
Отряд †Мезонихии (Mesonychia)
Семейство †Гапалодектовые (Hapalodectidae)
Род †Гапалодекты (Hapalodectes)
Вид †Hapalodectes lopatini
Вид †Hapalodectes paleocenus
Вид †Hapalodectes dux
Вид †Hapalodectes huanghaiensis
Вид †Hapalodectes hetangensis
Вид †Hapalodectes serus
Вид †Hapalodectes auctus
Вид †Hapalodectes lushiensis
Вид †Hapalodectes leptognathus
Вид †Hapalodectes anthracinus
Гапалодекты (Hapalodectes Matthew, 1909) — род семейства гапалодектовых (Hapalodectidae Szalay et Gould, 1966), происходящий из среднего палеоцена - среднего эоцена Азии и раннего эоцена Северной Америки. Установлены 10 видов.

Портретная реконструкция гапалодекта (Hapalodectes hetangensis). Автор — Р. Евсеев.
Синонимы: Dissacus Cope, 1881 (частью); Lohooedon.
Этимология
Родовое наименование образовано словами hapalos — "мягкий, нежный" и dêktês — "тот, кто кусает", означая буквально "мягкокусающий".


Череп гапалодекта (Hapalodectes hetangensis) (образец IVPP V12385). Палеозоологический музей Китая, Пекин.
Строение и функции
Большинство находок гапалодектов сводится к черепам и их фрагментам, частям нижних челюстей и отдельным зубам, принадлежащих разновозрастным особям (причем лучше сохранились черепа молодых животных). Размерный диапазон найденных черепов колеблется от 5 (видимо, молодое животное) до 10 см. Азиатские виды уступали по величине американским. Более крупный из них — Hapalodectes leptognathus — достигал размеров современной лесной куницы (Martes martes).
Предполагается, что череп гапалодектов имел крупные относительно тела размеры. Первое, что бросается в глаза при сравнении с Mesonychidae — относительно больший объем мозгового и укороченность лицевого отделов. Морфологически череп гапалодектов больше всего напоминает черепа некоторых примитивных хищников, например Vulpavus из Miacidae. Он умеренно длинный и узкий, с невысокими сагиттальным и затылочным гребнями (у взрослых особей гребни развиты заметно сильнее, чем у подростков). Передний край глазницы полукруглый и находится над P4. Глазница сзади замкнута, а слезная кость практически не расширяется в лицевом отделе, так что лобная кость подходит к верхнечелюстной полностью, в отличие от Mesonychidae, у которых глазница разомкнута, а слезная кость обладает выраженным лицевым расширением. Слезные отверстия внутренние, небольших размеров. Носовая кость длинная, узкая и несколько расширяется назад — она менее широкая, чем у Mesonychidae. Предчелюстная кость мала и расположена на передней части морды, так что носовая кость соприкасается с верхнечелюстной на большей части лицевого отдела. За альвеолами верхних резцов имеется пара маленьких отверстий.
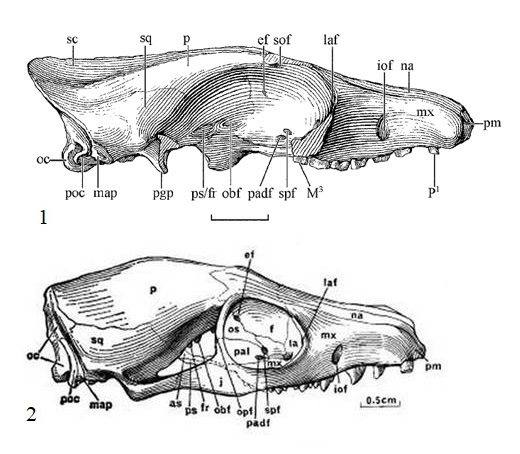
Прорисовки черепов гапалодекта (Hapalodectes hetangensis): 1 — взрослая особь; 2 — молодая особь с несменившимися зубами, типовой образец.
Верхнечелюстная кость короткая и высокая, с большим подглазничным отверстием над Р3. Последний находится на значительном расстоянии от переднего края глазницы, что свидетельствует об удлиненном инфраорбитальном канале. Скуловая кость относительно крупнее и массивнее, чем у представителей Miacidae, и очень похожа на таковую у Felidae. Лобная кость относительно узкая и слегка вогнутая между надглазничными гребнями. Заглазничный отросток лобной кости хорошо развит и соединяется с задней ветвью скуловой дуги. Чешуйчатая часть височной кости большая, узкая с боков и расширяется, придавая большую ширину задней части черепа. Скуловой отросток височной кости длинный, тонкий и соединяется со скуловой костью, накладываясь в косой шов. Теменная кость длинная, сравнительно узкая, выпуклая, в отличие от плоских теменных костей Mesonychidae, и охватывает большую часть черепной коробки.
Черепная коробка крупная, удлиненная, с боковым расширением в теменной части. Область основания черепа относительно больше, чем у Mesonychidae и скорее напоминает таковую Hyaenodonta и Miacidae. Затылочные мыщелки крупные, поперечно вытянутые и заметно разделены. Небная кость умеренно расширена в орбитальной области и не соприкасается со слезной костью. Суставные впадины поперечно расширены и ограничены четко фиксируемыми засуставным отростком сзади и низким гребнем впереди. Засуставное отверстие, отсутствующее у Mesonychidae, у гапалодектов хорошо выражено. У молодых особей слуховые пузыри слабо выражены либо полностью отсутствуют. Присутствуют 3 ветви внутренней сонной артерии среднего уха: мыса, стапедиальная и средняя ветвь внутренней сонной. Стременная ямка крупная.
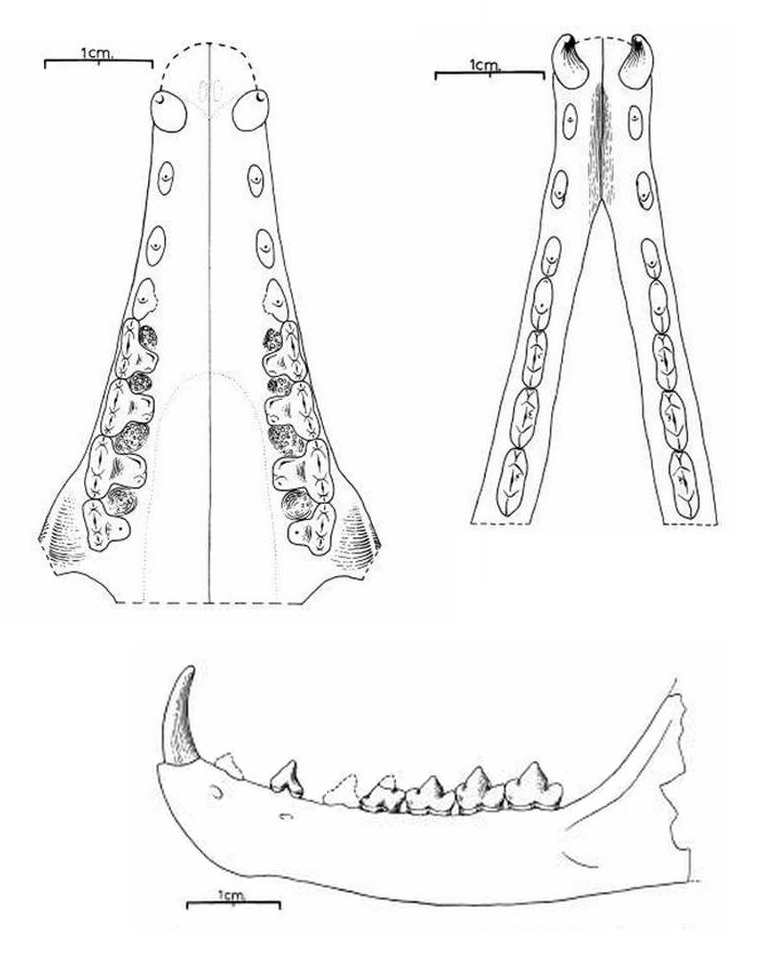
Прорисовки частично реконструированных верхней и нижней челюстей гапалодекта (Hapalodectes leptognathus).
Нижняя челюсть в целом выглядит довольно грацильно. Симфиз длинный и расширяется, начиная от переднего конца нижней челюсти до уровня ниже p2. Горизонтальная ветвь очень низкая и становится выше под m2-3. Венечный отросток большой и высокий. Суставный отросток крупный, поперечно вытянутый. Угловой отросток тонок и мал. Массетерная ямка большая, умеренно глубокая. Отмечается наличие 2 подбородочных отверстий под p1 и между p2-3 соответственно.
По строению зубов гапалодекты в общем схожи с другими мезонихиями. Наиболее важные отличия состоят в более сжатых с боков лезвиеподобных нижних щечных зубах, развитом гипоконе верхних коренных и входящих под углом выемках на нижних коренных. Верхние коренные имеют острые бугорки. Верхний клык большой и овальный в сечении, с заметным продольным гребнем с обеих сторон коронки. Нижний клык относительно большой, тонкий, и направлен немного вперед. К сожалению, отсутствуют находки, позволяющие четко определить количестве резцов. Предполагаемая зубная формула: I?3/i?3, C1/c1, P4/p4, M3/m3.
Распространение
Гапалодекты впервые появляются на территории Восточной Азии в среднепалеоценовое время. В Монголии и Китае их присутствие фиксируется до конца среднего эоцена. Спустя всего несколько миллионов лет, в самом начале эоцена они проникают в Северную Америку, где встречаются до конца раннеэоценовой эпохи. Вполне вероятно, что в раннем эоцене некоторые гапалодекты мигрировали из Северной Америки обратно в Азию, используя соединявшие в то время эти континенты сухопутные мосты.
Состав
Hapalodectes lopatini Solé, De Bast, Yang, Li et Smith, 2017 — наиболее древний вид, известный из среднего палеоцена (формация Доуму) Китая (провинция Аньхой). Описан по неполной нижней челюсти.

Неполная ветвь нижней челюсти гапалодекта (Hapalodectes lopatini).
Hapalodectes paleocenus Beard, Wang, Meng, Ni, Gebo et Li, 2010 — вид, известный из позднего палеоцена (формация Номоген, гашатский ярус) Китая (автономный район Внутренняя Монголия). Этот таксон описан по фрагменту левой ветви нижней челюсти с сохранившимися m2-3. Он крупнее, чем Hapalodectes hetangensis, но уступает по величине Hapalodectes leptognathus, Hapalodectes huanghaiensis и Hapalodectes serus. Как в размерном плане, так и по строению и пропорциям нижних коренных зубов к данному виду наиболее близок Hapalodectes dux, с которым он, вероятно, связан общим происхождением. Метакониды на нижних коренных выражены слабее, чем у Hapalodectes dux, Hapalodectes hetangensis и Hapalodectes huanghaiensis, но гораздо отчетливее, нежели у Hapalodectes anthracinus и Hapalodectes serus. m3 на 12 % крупнее m2. Массетерная ямка впереди распространяется до точки, условно расположенной под талонидом m3.
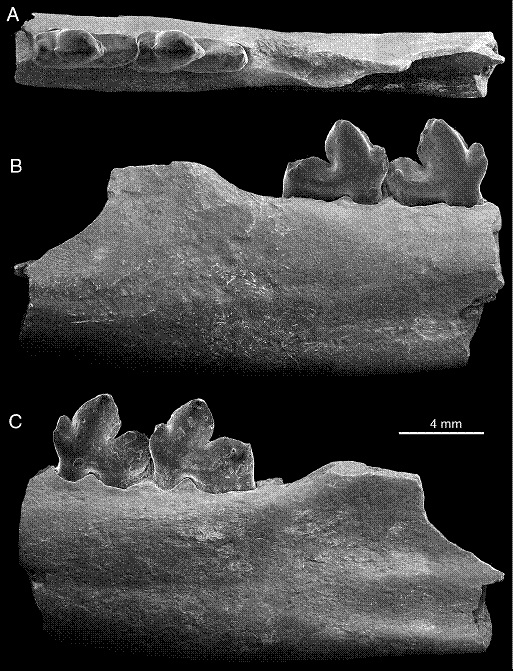
Фрагмент левой ветви нижней челюсти гапалодекта (Hapalodectes paleocenus) с m2-3; сверху (А), изнутри (В) и снаружи (С).
Hapalodectes dux Lopatin, 2001 — морфологически примитивный вид, известный из позднего палеоцена (формация Наран-Булак, гашатский ярус) Мрнголии (аймак Умнеговь). Описан по правой и левой ветвям нижней челюсти с большинством зубов. Данный вид мельче, чем Hapalodectes serus, но превосходит по размерам Hapalodectes hetangensis. Нижние коренные несут четко выраженные метакониды и протокристиды, а также рудиментарные гипоконулиды и энтокониды; m2 и m3 одинакового размера.
Hapalodectes huanghaiensis Tong et Wang, 2006 — вид, известный из раннего эоцена (формация Вуту, бумбанский ярус) Китая (провинция Шаньдун). Описан по серии зубов. На нижних коренных присутствуют четко выраженные метакониды. Вероятно, этот вид близок к Hapalodectes hetangensis. От Hapalodectes paleocenus его отличает наличие на талонидах нижних коренных 2 бугорков (вероятно, гипоконида и гипоконулида), которые повышаются в заднем направлении (а не ближе к середине, как у Hapalodectes paleocenus). Тригониды нижних коренных выше, острее и ориентированы более вертикально, чем у Hapalodectes paleocenus.
Hapalodectes hetangensis Ting et Li, 1987 — вид, известный из раннего эоцена (формация Лингча, бумбанский ярус) Китая (провинция Хэнань). Описан по полному черепу, ассоциированному с нижней челюстью с большей частью зубов, принадлежавших молодому животному. Также найден неполный череп взрослой особи с истертыми и отсутствующими зубами. По размерам этот вид явно меньше, чем североамериканский Hapalodectes leptognathus. Помимо величины, отличия от последнего состоят в отсутствии параконидов на P4, наличии 2 более развитых небольших бугорках под параконидами на нижних коренных, более развитым метаконидами нижних коренных и маленьким развитым бугорком на остром талониде m1-2. В отличие от более древних Hapalodectes dux и Hapalodectes paleocenus, у данного вида m3 несколько короче m2. От азиатского Hapalodectes serus этот вид отличается несколько меньшими размерами, а также более развитым метаконидами на нижних коренных, редуцированными парастилем и метастилем, поперечно более широкими и продольно более узкими верхними коренными. Хорошо развитые метакониды и наличие 2 небольших бугорков под параконидами нижних коренных указывают на большую примитивность по сравнению с Hapalodectes leptognathus и Hapalodectes serus.
Hapalodectes serus Matthew et Granger, 1925 — вид, известный из среднего эоцена (формация Аршанто, аршантский ярус - формация Шарамурун, шарамурунский ярус) Китая (автономный район Внутренняя Монголия). Описан по нижнему щечному зубу (p4 или m1). Этот узкий зуб не имеет метаконида, а удлиненный протоконид превратился в режущее лезвие. Позднее был найден фрагмент нижней челюсти со сломанными зубами. Отсутствие на нижних коренных метаконидов свидетельствует о уклонении рассматриваемого вида от основной линии развития гапалодектов. По данному признаку он может оказаться ближе к Hapalodectes anthracinus, чем к Hapalodectes hetangensis и Hapalodectes leptognathus. Вероятно, данное обстоятельство объясняется обменом фаунами между Северной Америкой и Азией во время раннего и среднего эоцена.
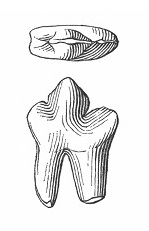
Прорисовки щечного зуба гапалодекта (Hapalodectes serus).
Hapalodectes auctus Matthew et Granger, 1925 — вид, известный из среднего эоцена (формация Ирдын-Манга, ирдынмангский ярус) Монголии. Описан по верхнему коренному зубу.
Hapalodectes lushiensis Chow, 1965 — поздний вид, известный из среднего эоцена (формация Луши) Китая (провинция Хэнань). Синоним: Lohooedon lushiensis. Описан по m3.
Hapalodectes leptognathus Osborn et Wortman, 1892 — типовой вид, известный из раннего эоцена (формации Уэрфано, Уосатч, Вайнд-Ривер и Виллвуд, уосатчский ярус) США (штаты Вайоминг и Колорадо). Синонимы: Dissacus leptognathus Osborn et Wortman, 1892 (тип); Hapalodectes compressus Matthew, 1909. Описан по фрагменту правой ветви нижней челюсти, в дальнейшем последовали другие относительно многочисленные находки. Крупнейший представитель рода. Как и у Hapalodectes paleocenus, метакониды на нижних коренных развиты слабо, но, в отличие от последнего, здесь имеются четко выраженные длинные паракониды, а гребни на талониде повышаются в заднем направлении.
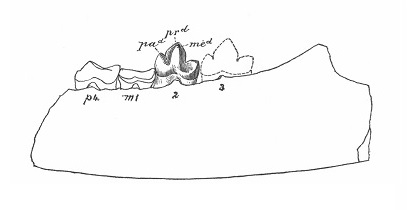
Прорисовка нижнечелюстного фрагмента гапалодекта (Hapalodectes leptognathus).
Hapalodectes anthracinus Zhou et Gingerich, 1991 — вид, известный из раннего эоцена (формация Виллвуд, уосатчский ярус) США (штат Вайоминг). Ископаемый материал очень фрагментарен и ограничивается изолированными зубами и фрагментами челюстей. Отличие от Hapalodectes leptognathus состоит в более коротких (на 15-16 %) нижних коренных с отсутствующими метаконидами. От азиатского Hapalodectes serus, помимо более коротких (на 11-12 %) нижних коренных, рассматриваемый вид отличается наличием на них более вертикальных, прямых и относительно высоких протоконидов. От Hapalodectes paleocenus он отличается рядом общих с Hapalodectes huanghaiensis характеристик зубного строения, в частности высокими, острыми и вертикально ориентированными тригонидами, а также повышающимися в заднем направлении гребнями талонидов нижних коренных. Однако общие размеры тела вполне сравнимы с Hapalodectes paleocenus.
Остатки, определенные лишь до рода и обозначенные Hapalodectes spp., известны из позднего палеоцена (формация Чицзян) Китая (провинция Цзянси), и раннего эоцена (формацииУосатч и Вайнд-Ривер, уосатчский ярус) и США (штат Вайоминг).
Образ жизни
Гапалодекты представляли собой небольших выдроподобных хищников. Считается, что они являлись преимущественно наземными, но не бегающими животными. Несомненно, гапалодекты вели активный хищный образ жизни. Их длинные, узкие и острые щечные зубы позволяют предполагать специализацию к рыбоедению.

Реконструкция предполагаемого образа жизни Hapalodectes.
Вымирание
Вероятно, окончательное исчезновение этих необычных мезонихий было связано с происходившей во второй половине эоцена глобальной перестройкой фауны, сопровождавшейся массовым вымиранием архаичных типов. К нему привела смена климата на более сухой и, как следствие, замещение тропических влажных лесов на обширные травянистые степи, саванны и редколесья.
Эволюционные связи
Краниодентальная морфология гапалодектов демонстрирует черты, отсутствующие у представителей семейства мезонихид (Mesonychidae): массивное основание черепа, замкнутые глазницы, весьма незначительное расширение слезной кости в лицевом отделе, заметное латеральное расширение черепной коробки (а значит, и увеличенный объем мозга), выдающийся гипокон верхних коренных, выемки на передней поверхности нижних коренных и др. В то же время, их нижние коренные зубы по большинству своих характеристик не отличаются от таковых у мезонихид. Все это дает возможность выделить гапалодектов и несколько родов, обладающими сходными с ними чертами, в отдельное семейство отряда мезонихий (Mesonychia) — гапалодектовых (Hapalodectidae).
Наличие сжатых с боков щечных зубов с продольно размещенными бугорками, мезиальных выемок на нижних коренных, небных ямок и некоторых других черт, сходных с таковыми у древних китов (Archaeoceti), позволили выдвинуть предположение о родстве гапалодектов (и гапалодектид в целом) с китообразными (Cetacea). Однако последние исследования показали, что зубы и черепа древних китов (например, Pakicetus, Protocetus и др.) больше напоминают таковые мезониховых. Исходя из данного критерия, китообразные оказываются ближе к мезонихидам, а гапалодектиды состоят с последними в более дальнем родстве, чем предполагалось ранее.
Ссылки
Гапалодекты (Википедия) перейти
Hapalodectes (Paleobiology Database) перейти
Литература
Зоогеография палеогена Азии / Отв. ред. Крамаренко Н. Н. — М.: Наука, 1974
Кэрролл Р. Палеонтология и эволюция позвоночных: В 3-х т. Т. 2. — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 13: Млекопитающие / Гл. ред. Орлов Ю. А. — М.: Государственное научно-техническое издательство литературы по геологии и охране недр, 1962
Beard K. C., Wang Y.-Q., Meng J., Ni X.-J., Gebo D. L., Li C.-K. Paleocene Hapalodectes (Mammalia: Mesonychia) from Subeng, Nei Mongol: further evidence of "East of Eden" dispersal at the Paleocene-Eocene boundary. Vertebrata PalAsiatica 48 (4), 2010
Macdonald D. W. The Velvet Claw. A Natural History of the Carnivores. — New York: Parkwest; London: BBC Books, 1993
O'Leary M. A. Morphology of the humerus of Hapalodectes (Mammalia, Mesonychia). American Museum novitates 3242, 1998
Rose K. D. The Beginning of the Age of Mammals. — Oxford: Johns Hopkins University Press, 2006
Szalay F. S. The Hapalodectine and a phylogeny of the Mesonychidae (Mammalia, Condylarthra). American Museum novitates 2361, 1969
Szalay F. S., Gould S. J. Asiatic Mesonychidae (Mammalia, Condylarthra). Bulletin of the American Museum of Natural History 132, 1966
Ting S., Li C. The skull of Hapalodectes (?Acreodi, Mammalia), with notes on some Chinese Paleocene mesonychids. Vertebrata PalAsiatica 25, 1987
Ting S., Wang Y., Schiebout J. A., Koch P. L., Clyde W. C., Bowen G. J., Wang Y. New Early Eocene mammalian fossils from the Hengyang Basin, Hunan China. Bulletin of Carnegie Museum of Natural History 36, 2004
Zhou X., Gingerich P. D. New species of Hapalodectes (Mammalia, Mesonychia) from the Early Wasatchian, Early Eocene, of northwestern Wyoming. Contributions from the Museum of Paleontology, University of Michigan 28, 1991