Гемипсалодоны — Hemipsalodon
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Клада Скротиферы (Scrotifera)
Клада Феревунгуляты (Fereuungulata)
Клада Ферэ (Ferae)
Клада Всехищные (Pan-Carnivora)
Отряд †Гиенодонты (Hyaenodonta)
Надсемейство †Гиайнайлюроиды (Hyainailouroidea)
Семейство †Гиайнайлюровые (Hyainailouridae)
Подсемейство †Гиайнайлюрины (Hyainailourinae)
Род †Гемипсалодоны (Hemipsalodon)
Вид †Hemipsalodon viejaensis
Вид †Hemipsalodon grandis
Гемипсалодоны (Hemipsalodon Cope, 1885) — род подсемейства гиайнайлюрин (Hyainailourinae (Pilgrim, 1932)), происходящий из среднего - позднего эоцена Северной Америки. Установлены 2 вида.

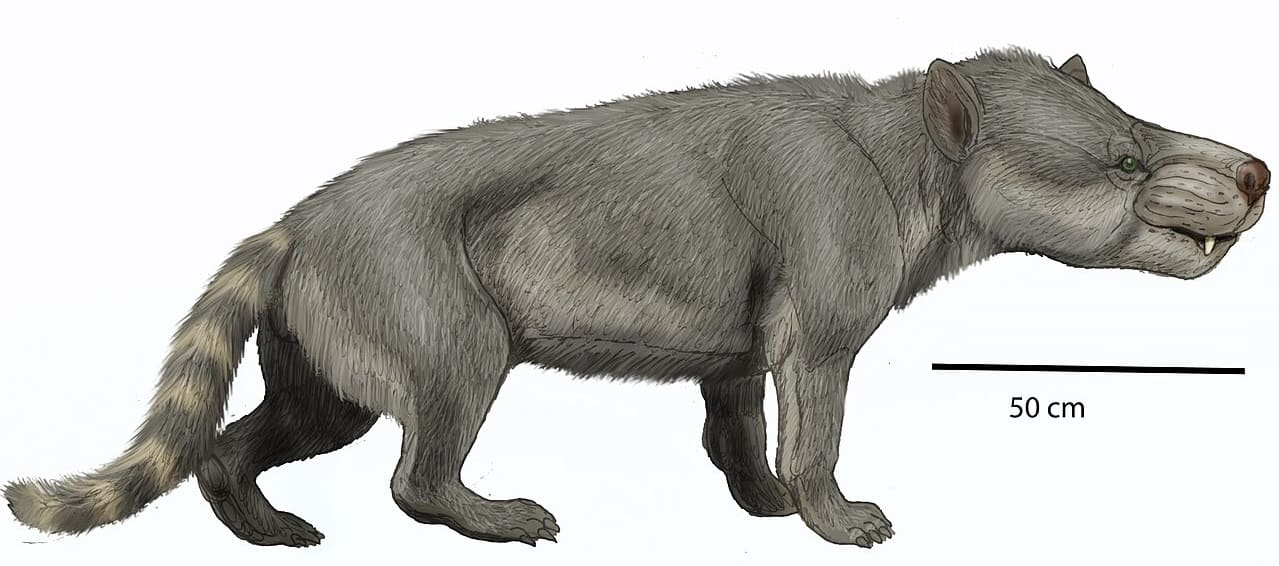
Реконструкция внешнего облика гемипсалодона (Hemipsalodon grandis). Автор — Р. Евсеев.
История изучения
Род и вид Hemipsalodon grandis были описаны Копом (1885 г.) на основе фрагментов нижней челюсти и бедренной кости, обнаруженных в отложениях формации Сайпресс-Хиллз на крайнем юге канадской провинции Саскачевана. Находки имели позднеэоценовый (чадронский) возраст и, по мнению автора открытия, принадлежали оксиениду. Некоторое время спустя Ламб (1908 г.) сообщил о найденном там же крупном фрагменте клыка, также относящегося к Hemipsalodon.
Расселл (1934 г.) в своем обзоре фауны Сайпресс-Хиллз сообщил о верхнем зубном ряде Hemipsalodon grandis, а позднее (1938 г.) описал переднюю часть его черепа и предпринял попытку реконструировать этого крупного хищника. Он верно изобразил крупное медведеподобное животное, но недооценил общей длины и ширины черепа.

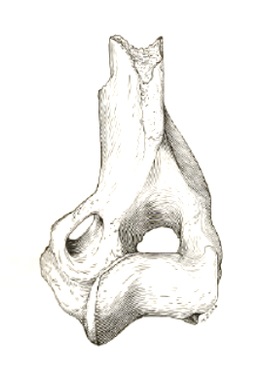
Прорисовки скелетных остатков гемипсалодона (Hemipsalodon grandis) из позднего эоцена штата Вайоминг, США: слева — фрагмент левой ветви нижней челюсти с p2-m1, сверху и снаружи; справа — дистальная часть левой плечевой кости, спереди.
Шлейкджер (1935 г.) обнаружил фрагмент нижней челюсти и часть плечевой кости в позднеэоценовых слоях Йодер формации Чадрон на юго-востока Вайоминга, США, и описал на их основе новый вид, названный им Hemipsalodon cooki. Позже, однако, он был признан младшим синонимом Hemipsalodon grandis. Вероятно, типовому виду принадлежат и остатки, описанные в качестве Hemipsalodon sp. из чадрония формации Уайт-Ривер в Вайоминге, США.

Череп с нижней челюстью гемипсалодона (Hemipsalodon grandis) из среднего эоцена штата Орегон, США.
Эвернден и др. (1964 г.), основываясь на идентификации фоссилий Мак-Кенной, сообщили о находках ископаемых остатков Hemipsalodon grandis в среднеэоценовых (дюшенских) отложениях формации Кларно в центральной части Орегона, США. Помимо этого, фрагментарные остатки Hemipsalodon grandis были обнаружены в среднем эоцене (дюшении) формации Галистео на территории Нью-Мексико, США.
Второй установленный на сегодняшний день вид, Hemipsalodon viejaensis, был описан Густафсоном (1986 г.) по находкам зубов P4, M1 и M2 из среднеэоценовых (дюшенских) отложений формации Чамберз-Тафф в Техасе, США. Таким образом, ареал обитания рода простирался довольно широко — от юга Канады до границы США с Мексикой.
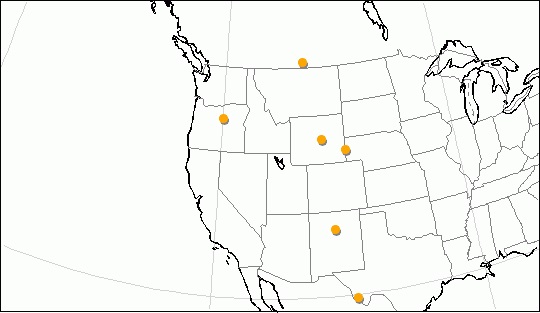
Карта местонахождений ископаемых остатков Hemipsalodon.
Строение и функции
При черепе длиной в 45 см и расчетной длине тела 180 см (без учета хвоста) гемипсалодоны являются крупнейшими североамериканскими гиенодонтами; вероятно, их масса могла достигать 200 кг. Хотя находки посткраниального скелета весьма фрагментарны, с определенной степенью уверенности можно заключить, что животные в общих чертах напоминали медведя с вытянутым телом. Находка астрагала позволяет предположить, что конечности были стопоходящими.


Череп с нижней челюстью гемипсалодона (Hemipsalodon grandis) из среднего эоцена штата Орегон, США.
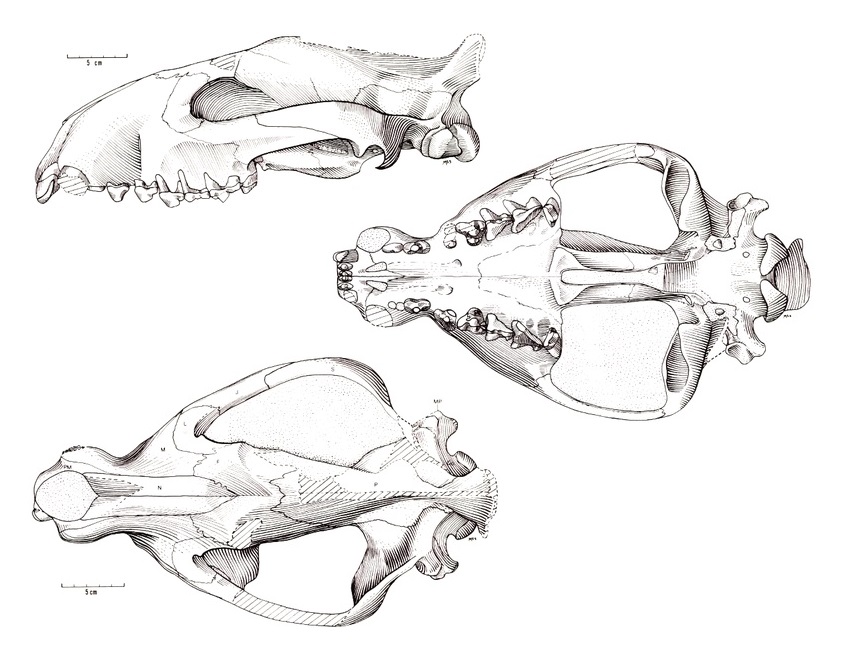
Череп гемипсалодонов внешне очень массивный, при виде сбоку продолговатый, с сильными и широкими горизонтальными скуловыми дугами, заметно расширяющими череп латерально. Носовое отверстие довольно большое и передняя часть морды с массивными и длинными клыками выглядит приподнятой кверху. Лобная область черепа широкая спереди, но значительно сужается назад. Массивные скуловые кости свидетельствуют о хорошо развитом комплексе жевательной мускулатуры, напоминающем скорее таковой типичных представителей отряда Oxyaenodonta. Гребешок, выходящий из задней части скуловой дуги, соединяется с сосцевидным отростком, сильно его расширяя. Засуставный отросток массивен и пронизан большим засуставным отверстием. Черепная коробка весьма мала. Сагиттальный гребень не сохранился, но, судя по всему, был достаточно высоким. Затылочный гребень высокий и узкий, при виде сзади затылочная область черепа имеет форму песочных часов.

Прорисовки реконструированных черепа и нижней челюсти гемипсалодона (Hemipsalodon grandis).
Небо достаточно широкое и вогнуто во фронтальной плоскости. Вдоль средней линии неба развит сильный гребень, который соединяется с занебным валиком, расположенным поперечно, сразу позади линии M3. Наблюдается удлинение задней части неба, но без развития в дыхательную костную трубку, характерную для Hyaenodon. Основание черепа умеренно укорочено, но не настолько сжато в переднее-заднем направлении, как у крупных видов Hyaenodon. К сожалению, детали строения базикраниума не сохранились.
Нижняя челюсть довольно высокая, заметно выше, чем у Pterodon africanus. Симфиз складчатый и наклонен вперед примерно на 45 °. 2 подбородочных отверстия расположены на боковой стороне челюсти: одно под p1, другое немного впереди соприкосновения p3 и p4. Большая часть венечного отростка не сохранилась, однако некоторые детали различимы. Передний край венечного отростка расширен латерально до формы гребня. Массетерная ямка неглубокая и расположена по боковой стороне челюсти. Нижняя часть ямки постепенно наклоняется на основной части ветви. Постеровентральная часть ямки образует небольшой гребень, поднимающийся дорсально и латерально на широкий и массивный сустав.
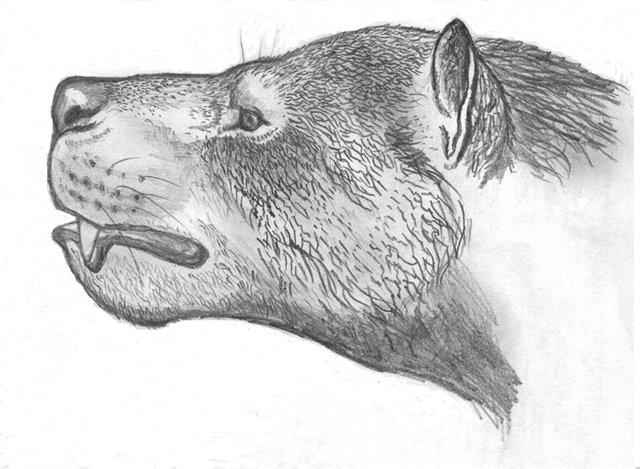
Портретная реконструкция гемипсалодона (Hemipsalodon grandis). Автор — А. Гордеев.
Зубная формула гемипсалодонов составляет I3/i3, C1/c1, P4/p4, M3/m3. Зубная морфология отличается от Hyaenodon наличием M3 и отсутствием проксимодистально ориентированных режущих граней на Ml-2/m2-3; отличая от Pterodon состоят в гораздо более крупных размерах зубов, широко разделенном параконом и метаконом М2, имеющем относительно более крупные размеры M3, отчетливых воротничках на лингвальной поверхности верхних и нижних зубов.
Пожалуй, наиболее интересной особенностью строения зубной системы гемипсалодонов является проворачивание верхних и нижних коренных зубов. И те, и другие повернуты по проксимодистальным зубным осям: верхние — медиально (вовнутрь), нижние — латерально (наружу). Это означает, что на протяжении жизни животного верхние и нижние коренные, постоянно наклоняясь друг к другу, тесно соприкасались и самозатачивались. Практически, это позволяло им до самой старости иметь достаточно острые зубы. Верхний зубной ряд демонстрирует аналогичный характер износа и подтверждает предположение о проворачивании коренных зубов. Первоначально считалось, что такое проворачивание стало результатом посмертных изменений. Однако если зубы восстановить в вертикальной позиции в челюсти, стертые грани на верхних и нижних коренных располагаются так, что прикус практически невозможен, и выпадает мыщелок из суставной ямки. Необходимо отметить, что подобное проворачивание коренных в онтогенезе наблюдается и у представителей рода Hyaenodon, однако не замечено ни у кого из Hyainailourinae — подсемейства, в котором состоит Hemipsalodon.

Портретная реконструкция гемипсалодона (Hemipsalodon grandis). Автор — Р. Евсеев.
Верхние резцы расположены в слегка выпуклый ряд. I3 больше, чем 2 других резца, приблизительно равных между собой по размеру. Большая диастема между I3 и большим верхним клыком служила для вкладывания массивного нижнего клыка. Верхний клык эллиптический в сечении, со значительной в проксимодистальной оси шириной. У молодых особей эмаль клыка обладает складчатой фактурой, и крупные борозды эмали спускаются к дистально-язычной границе зуба.
P1 и P2 имеют двойные корни. К сожалению, они не сохранились на черепе из формации Кларно и мало что можно сказать о них. P3 обладает небольшим передним и крупным задним корнем. По центру коронки расположена крупная вершинка, однако о деталях ее строения трудно судить из-за плохой сохранности коронки. P4 напоминает таковой у Hyaenodon — это большой, имеющий 3 корня зуб, с массивным воротничком, опоясывающим коронку. Основная лабиальная вершинка высокая, сжатая с боков и занимает центральное положение. Передний гребень проходит от вершинки к воротничку; задний гребень спускается с вершинки, но прерывается большим задним базальным выступом. Судя по размерам медиального корня, протокон является важным острием.

Портретная реконструкция гемипсалодона (Hemipsalodon viejaensis). Автор — Р. Евсеев.
М1 — треугольный зуб с различимыми параконом, метаконом и протоконом. Метакон ориентирован проксимолингвально по отношению к паракону, обе вершинки сжаты с боков. Сильный, лезвиеподобный метастиль продлевается околощечно из задней части зуба. M2 в основном похож на М1, за исключением того, что он намного больше, а паракон и метакон более сжаты. Протокон мал и направляет препаракристу и постпаракристу в щечном направлении к основанию паракона и метакона соответственно. M3 имеет форму равнобедренного треугольника с широким основанием. Один большой параметакон находится практически в центре зуба. Различные препаракристы пролегают от этой вершинки дистобуккально к краю M3. Постметакриста также наблюдается, но произвести оценку степени ее развитости затрудняет плохая сохранность. Протокон намного меньше, чем параметакон и направляет препротокристу в щечном направлении, которая объединяет его с основанием параметакона. Постпротокриста пролегает проксимобуккально от протокона к краю зуба, но повреждение зуба слишком обширное, чтобы различить прочие детали. Наблюдаются глубокие впадины в верхней челюсти между M1-2 и М2-3 для вкладывания больших тригонидов m2-3.

Портретные реконструкции гемипсалодонов — Hemipsalodon viejaensis (на переднем плане) и Hemipsalodon grandis. Автор — Р. Евсеев.
Нижний клык большой и покрыт толстым слоем эмали. p1 обладает 1 корнем и слегка повернут буккально, относительно основной оси зубного ряда; коронка не сохранилась. p2 имеет 2 корня, однако его коронка также не сохранилась. p3 умеренно высокий и относительно длинный, с основной вершинкой, оснащенной передним и задним гребнями, спускающихся от вершинки к основанию зуба. Маленькие бугорки располагаются в основании заднего гребня. На лингвальной стороне зуба различим воротничок. p4 большой и массивный, напоминает аналогичный у Hyaenodon. Маленькие бугорки расположены на дистолингвальном углу зуба, поднимаясь на высокую основную вершинку и опускаясь сзади на большую заднюю базальную вершинку.
m1 той же длины, что и p4, однако гораздо более узок букколингвально и развивается как плотоядный зуб. Как и у Hyaenodon, m1, вероятно, окклюзирует с хищническим dP4. К сожалению, отсутствуют образцы с целым m1 и, следовательно, мало что можно добавить к этому описанию. m2 больше m1 и оснащен большим лезвием, состоящим из расширенного паракристида. Протоконид выше, чем параконид, и нет никаких следов метаконида. Режущие грани также имеет место на задней части протокристида. Развит довольно большой талонид. m3 является самым крупным из нижнего ряда коренных зубов и похож на m2 во всех отношениях, за исключением немного меньшего талонида. Хотя плохая сохранность затрудняет судить уверенно о структуре зуба, кажется, нет никаких признаков дистолингвального усиления на преваллиде M2 и M3, как у Pterodon.

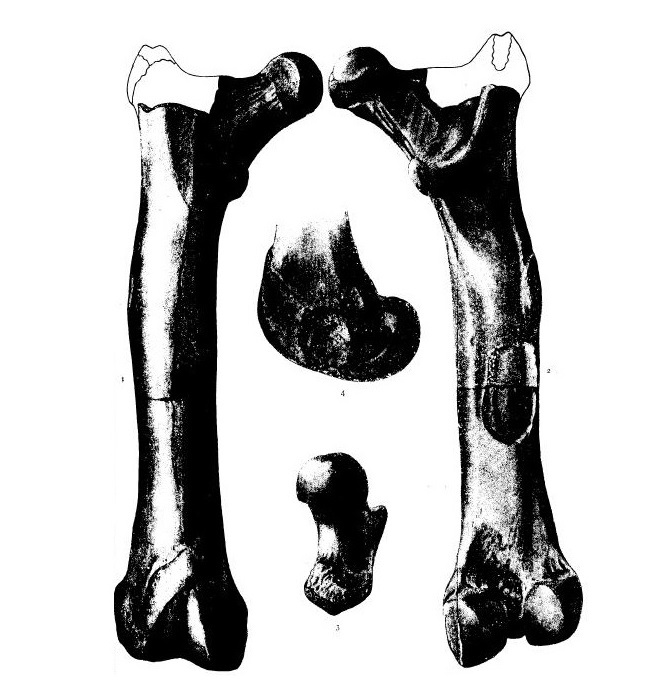
Бедренная кость гемипсалодона (Hemipsalodon grandis) из позднего эоцена провинции Саскачеван, Канада, и ее прорисовки.
Особого упоминания заслуживает находка очень крупных ископаемых остатков, формально причисляемых Hemipsalodon, но вряд ли им принадлежащих. Она представляет собой 2 парные плечевые кости длиной более 50 см, принадлежащие какому-то чрезвычайно крупному хищнику. Они прямые и толстые и превышают по размерам плечевые кости крупного быка. Расчетная длина черепа их обладателя составляет не менее 70 см в длину. Кому эти кости принадлежали на самом деле — пока неизвестно. Наиболее вероятно, что их обладателями являлись какие-то крупные мезонихии (Mesonychia).
Реконструкция внешнего облика немипсалодона (Hemipsalodon grandis). Автор — Д. Богданов.
Состав
Hemipsalodon viejaensis Gustafson, 1986 — вид, известный из среднего эоцена (формация Чамберз-Тафф, дюшенский век) штата Техас, США. Более мелкий представитель рода: длина черепа составляет 40 см.
Прорисовки черепа гемипсалодона (Hemipsalodon viejaensis) из среднего эоцена штата Техас, США.
Hemipsalodon grandis Cope, 1885 (тип) — типовой вид, известный из среднего - позднего эоцена (формации Кларно, Галистео и Чадрон, дюшенский - чадронский века) штатов Орегон, Вайоминг и Нью-Мексико, США, а также позднего эоцена (формация Сайпресс-Хиллз, чадронский век) провинции Саскачеван, Канада. Синоним: Hemipsalodon cooki Schlaikjer, 1935. Череп достигает 45 см в длину.


Череп и нижняя челюсть гемипсалодона (Hemipsalodon grandis) из среднего эоцена штата Орегон, США.
Остатки, определенные лишь до рода и обозначенные Hemipsalodon sp., известны из позднего эоцена (формация Уайт-Ривер, чадронский век) штата Вайоминг, США.

Гемипсалодон (Hemipsalodon grandis) в естественном окружении. Автор — Р. Евсеев.
Образ жизни
Период существования гемипсалодонов пришелся на время, когда еще распространялись теплые тропические и субтропические зоны. Во влажных лесах обитало множество растительноядных животных, обеспечивавших этих хищников пищей. Гемипсалодоны могли охотиться на слабых особей, детенышей, питаться падалью. Возможно, что они, подобно современным медведям, отбирали добычу у меньших по размеру хищников, например у ранних представителей отряда Carnivora (Nimravidae, Amphicyonidae) и у других гиенодонтов (Hyaenodon и т.п.), подбирали остатки их трапезы.

Кормящийся гемипсалодон (Hemipsalodon grandis).
Вымирание
В конце эоцена в относительно короткие сроки планета пережила глобальное изменение климата. Три события — резкое похолодание в Северной Америке, установление засушливого климата в Азии и образование ледяного щита Антарктиды — произошли практически одновременно в течение нескольких сотен тысячелетий. Повсеместное сокращение тропиков и их замещение более сухими и прохладными умеренными зонами повлекло за собой массовое вымирание фитофагов, специализированных на питании мягкой теплолюбивой растительностью, а следом за ними — многих специализированных хищников, включая и гемипсалодонов.
Ссылки
Hemipsalodon (Paleobiology Database) перейти
Литература
Лавров А. В. Неизвестные хищники креодонты. ПалеоМир 1 (8), 2010
Cope E. D. The Vertebrata of the Tertiary and Cretaceous Rocks of the North West Territory. 1891
Evolution of Tertiary Mammals of North America. Vol. 1: Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals. Edited by Janis C. M., Scott K. M. & Jacobs L. L. — Cambridge University Press: Cambridge, 1998
Prothero D. R., Emry R. J. The terrestrial Eocene-Oligocene transition in North America. — Cambridge University Press: Cambridge, 1994
Silvan M. J. A skull of Hemipsalodon (Mammalia, Deltatheridia) from the Clarno Formation of Oregon. American Museum novitates 2387, 1969