Гобиатерии — Gobiatherium
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Клада Скротиферы (Scrotifera)
Клада Феревунгуляты (Fereuungulata)
Клада Эвунгуляты (Euungulata)
Клада Мезаксонии (Mesaxonia)
Отряд †Диноцераты (Dinocerata)
Семейство †Гобиатериевые (Gobiatheriidae)
Род †Гобиатерии (Gobiatherium)
Вид †Gobiatherium mirificum
Гобиатерии (Gobiatherium Osborn et Granger, 1932) — монотипический род монотипического семейства гобиатериевых (Gobiatheriidae Flerov, 1952), происходящий из раннего - среднего эоцена Азии.
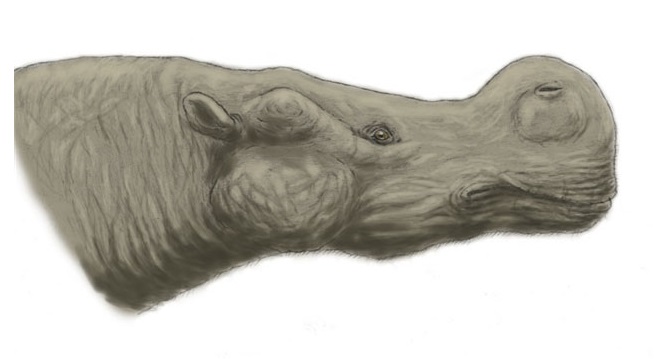
Реконструкция внешнего облика гобиатерия (Gobiatherium mirificum). Автор — Д. Богданов.
Единственный установленный вид — Gobiatherium mirificum Osborn et Granger, 1932 (тип). Его остатки обнаружены в Китае (автономный район Внутренняя Монголия, провинции Хубэй, Хенань и Сычуань), Казахстане и Киргизии. Они датируются ранним - средним эоценом (аршантский - ирдынмангский ярусы). Синонимы: Gobiatherium major Qi, 1987; Gobiatherium monolobotum Qi, 1987; Gobiatherium minutum Cheng et Ma, 1990.
История изучения
Родовое название
Gobiatherium означает "зверь из Гоби" и было дано по первому месту обнаружения ископаемых остатков. Череп и нижняя челюсть данного животного описали в 1932 г. Генри Ферфилд Осборн и Уолтер Виллис Грейнджер из местонахождения, расположенного в 40 км к югу-западу от Ирен-Дабасу во Внутренней Монголии, где их извлекли из среднеэоценовых (ирдынмангских) отложений формации Ирдын-Манга. Здесь были найдены как минимум 7 черепов различной степени сохранности, включая полный типовой череп, ювенильная нижняя челюсть и элементы посткраниального скелета. Позже нижняя челюсть была обнаружена в Улан-Шире, а отдельные зубы — в Сычуане.
Исследователи с черепом гобиатерия (Gobiatherium mirificum) из среднего эоцена (ирдынмангия) автономного района Внутренняя Монголия, Китай.
Почти полный череп был выкопан в 1971 г. из среднеэоценовых отложений формации Чонкурчак у озера Иссык-Куль в Киргизии. В 1983 г. разрозненные остатки гобиатерия извлекли из среднэоценовых (аршантских) отложений формации Чакпактас в Казахстане. Gobiatherium minutum, описанный по фрагментарным остаткам из раннеэоценовых отложений формации Юйхуандин из провинции Хубэй, Китай, скорее всего, является младшим синонимом типового вида.
Неполный череп с нижней челюстью гобиатерия (Gobiatherium mirificum) из среднего эоцена (ирдынмангия) автономного района Внутренняя Монголия, Китай.
Родственные связи
Gobiatherium является диноцератом, обладающим множеством специфических черт. У него отсутствуют рогообразные выросты и бивни, характерные для большинства родственников, но имеются широко расставленные скуловые дуги и расширенная, аркообразно поднятая носовая область. Первоначально Gobiatherium причислили к семейству Uintatheriidae, однако Константин Константинович Флеров в 1952 г. выделил этот род в самостоятельное семейство Gobiatheriidae. Уолтер Х. Уилер в 1961 г. пересмотрел его статус в качестве подсемейства Gobiatheriinae в составе Uintatheriidae, с чем согласились Роберт Милтон Шох и Спенсер Джордж Лукас в 1985 г. На сегодняшний день систематическое положение гобиатерия остается предметом дискуссий.
Неполный череп гобиатерия (Gobiatherium mirificum) из среднего эоцена Тору-Айгыр, Киргизия.
Происхождение
Gobiatherium также остается невыясненным. Представители более древних родов, таких как
Prodinoceras или
Bathyopsis, значительно отличаются своей черепной морфологии и, следовательно, напрямую с ним не связаны. В строении посткраниального скелета гобиатерий обладает отчетливым сходством с Uintatheriidae — прогрессивными диноцератами, потомками архаичных Prodinoceratidae. Остатки Uintatheriidae долгое время были известны только из Северной Америки. Бытовала точка зрения, согласно которой от их американской линии произошли животные, которые проникли в Азию и стали предками
Gobiatherium. Однако затем в Китае были обнаружены остатки представителей рода
Uintatherium, датированные ранним и средним эоценом. Неизвестно, были ли это потомки американских диноцерат, или, что более вероятно, азиатских. Таким образом, нельзя исключать и "местное", азиатское происхождение
Gobiatherium.
Череп гобиатерия (
Gobiatherium mirificum) из среднего эоцена (аршантия) Китая.
Строение и функции
Гобиатерий представляет собой довольно крупного диноцерата: его типовой череп достигает 68 см в длину, расчетная высота в холке составляет около 1,25 м, общая длина тела — 2,8 м, а вес оценивается в 1,5 т.
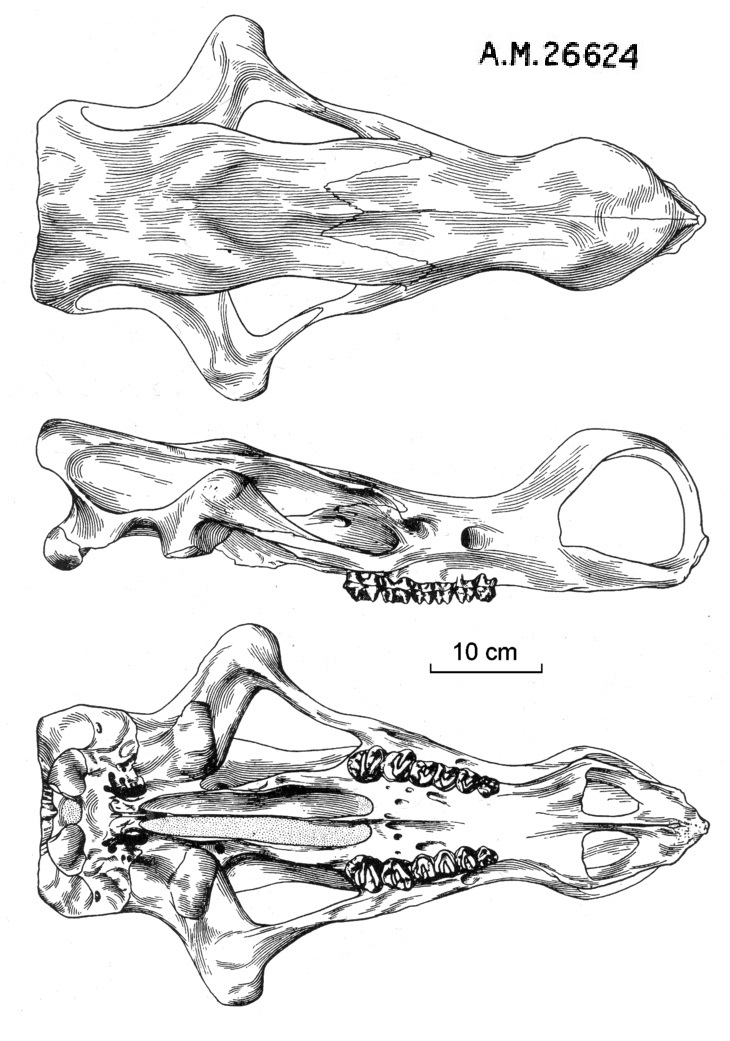
По своему строению череп резко отличается от черепов других диноцерат, что, очевидно, указывает на высокую специализацию. Череп удлиненный, очень низкий в лобной и затылочной областях, без рогообразных выростов на верхнечелюстных и теменных костях. Расширенная носовая область высоко аркообразно поднята посередине, иногда на ней присутствует пара маленьких выростов, напоминающих рожки — вероятно, проявление полового диморфизма, так же как и костные дуги, соединяющие концы носовых костей с вершинами предчелюстных и укрепляющие носовую область. Скуловые дуги очень широко раздвинуты в стороны, резко изогнуты в области сочленовных впадин. Сагиттальный гребень отсутствует. Теменные гребни широко раздвинуты, пространство между ними плоское, за исключением небольшого вздутия на лбу посередине между глазницами и затылком. Затылок четырехугольный, с выступающими мыщелками.
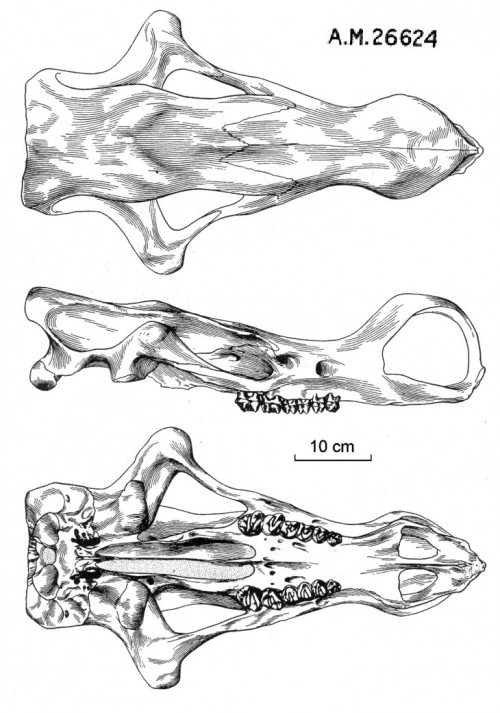
Прорисовки черепа гобиатерия (Gobiatherium mirificum) из среднего эоцена (ирдынмангия) автономного района Внутренняя Монголия, Китай; сверху, сбоку и снизу.
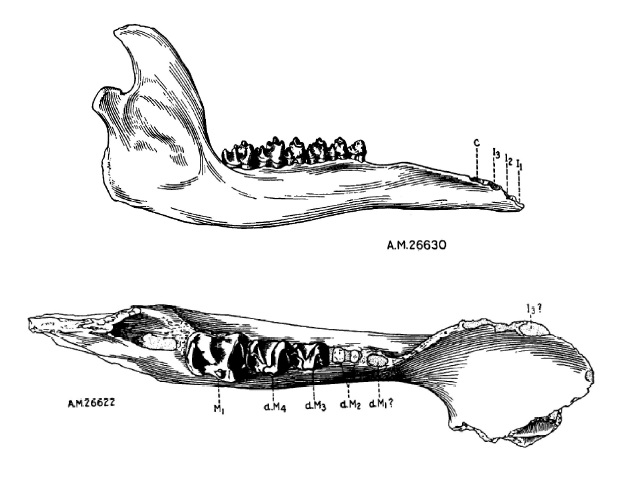
Небная хоанальная вырезка с большим средним отростком, далеко выступающим назад. Сошник полностью окостеневший и полностью разделяет задний хоанальный вход надвое, опираясь на задний средний отросток небных костей, далеко выступающий назад. Задние концы небных костей, ограничивающие с боков хоану, толсты, но не образуют гребня, окаймляющего хоанальный вход. Нижняя челюсть в области симфиза лопатообразная, неглубокая и широкая. Ее ветви поднимаются высоко (выше, чем у Uintatherium). Предохранительные лопасти совершенно не развиты в связи с отсутствием верхних клыков. По всей видимости, были развиты мясистые губы.
Прорисовки нижних челюстей гобиатерия (Gobiatherium mirificum) из среднего эоцена (ирдынмангия) автономного района Внутренняя Монголия, Китай; сбоку и сверху.
Зубная формула: I0/i3, C0/c1, P3/p3, M3/m3 = 32. Верхние резцы и клыки отсутствуют. У одной особи выявлен рудиментарный клык с одной стороны верхней челюсти. Нижние резцы двухлопастные. Нижние клыки небольшие, предположительно низкие, резцеобразные. Предкоренные и коренные зубы в общих чертах сходны с таковыми у Uintatheriidae. Предкоренные достигают почти полной моляризации и раскрытия долинки. р1, возможно, имеется у молодых (молочный), у взрослых отсутствует. В отличие от Uintatheriidae, М3 не приобретает дополнительной вершинки, т. е. зуб не усложняется — это свидетельствует о том, что пищевой режим не изменился в сторону включения более твердых элементов. Щечные зубы образуют при смыкании плотный замок, исключающий какие-либо боковые и передне-задние движения челюстей. Зубочелюстной аппарат, безусловно, приспособлен к потреблению мягких кормов.
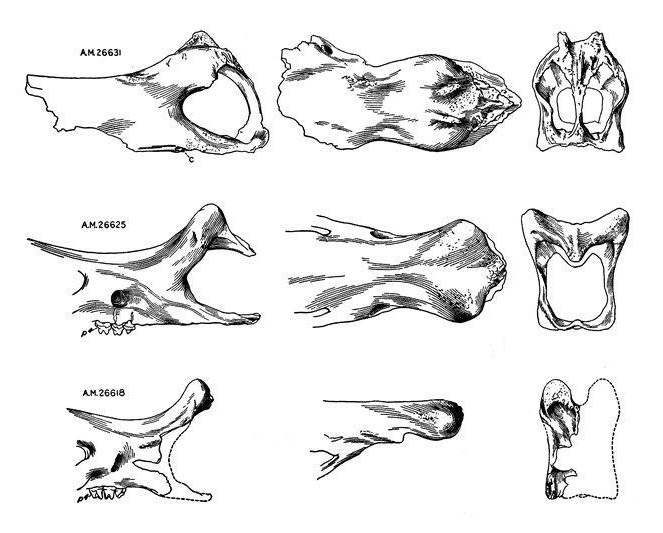
Различия в строении носовых костей трех черепов гобиатерия (Gobiatherium mirificum) из среднего эоцена (ирдынмангия) автономного района Внутренняя Монголия, Китай: вверху и посередине — взрослые животные, внизу — ювенильная особь. Изображения даны в одном масштабе.
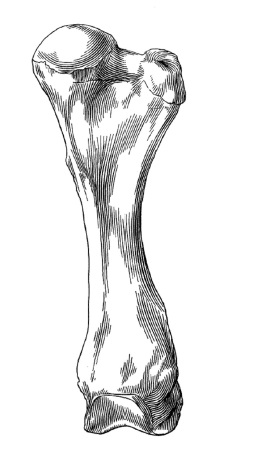
Посткраниальный скелет известен лишь частично. Конечности относительно длинные. Плечо и бедро напоминают таковые Uintatherium. Бедро без III-го вертела. Пястные и плюсневые кости удлиненные и тонкие. В заплюсне II-я и III-я клиновидные кости раздельны, астрагал без шейки. Задняя конечность пальцеходящая, передняя — неизвестно.
Прорисовка левого бедра гобиатерия (Gobiatherium mirificum) из среднего эоцена (ирдынмангия) автономного района Внутренняя Монголия, Китай; спереди.
Образ жизни
Морфологические особенности гобиатерия указывают на его полуводный образ жизни, сходный с таковым гиппопотамов (
Hippopotamus). В строении черепа это выражается в высоко поднятых носовых костях и широком нижнечелюстном симфизе. Аркообразно возносящийся носовой отдел черепа, вероятно, служил для облегчения дыхания при погружении морды в воду во время добывания пищи, ноздри же при этом оставались над водной поверхностью, подобно таковым у эмболотериев (
Embolotherium). В пользу этого говорит и то, что известные черепа, различающиеся деталями строения, вероятно, отличающими самцов от самок, все имеют одинаково поднятый носовой отдел. Лопатообразно расширяющаяся кпереди нижняя челюсть с широкими двухлопастными резцами, по-видимому, действовала наподобие челюсти платибелодонов (
Рlatybelodon) — захватывала и вытаскивала из воды растительные массы.
Портретная реконструкция гобиатерия (
Gobiatherium mirificum). Авторы — А. Молош, Р. Евсеев.
Низкий череп и простиравшееся далеко назад твердое небо, в целом свойственные водным животным, также подтверждают предположение о амфибиотическом образе жизни гобиатерия. Вероятно, именно в связи с этим животное обладало удлиненными и тонкими метаподиями, а пальцы его кисти и стопы были способны широко раздвигаться, как у бегемотов, создавая большую площадь опоры на топкой почве. Судя по всему, гобиатерий держался по берегам рек и озер, где питался сочной водной растительностью.
Гобиатерий (
Gobiatherium mirificum) в естественном окружении. Автор — Н. Комаров.
Вымирание
Гобиатерий дожил почти до конца среднего эоцена, он был одним из последних диноцерат. Возможно, именно высокая специализация позволила ему просуществовать дольше большинства других представителей отряда. Представляется вероятным, что гобиатерий исчез в связи с неблагоприятными изменениями климата, повлекшими за собой иссушение его местообитаний.
Ссылки
Gobiatherium (Paleobiology Database)
перейти
Литература
Зоогеография палеогена Азии / Отв. ред. Крамаренко Н. Н. — М.: Наука, 1974
Молош А. Альтернативные звери раннего палеогена. В мире животных 2, 2012
Основы палеонтологии. В 15 т. Т. 13: Млекопитающие / Гл. ред. Орлов Ю. А. — М.: Государственное научно-техническое издательство литературы по геологии и охране недр, 1962
Флеров К. К. Диноцераты Монголии. Труды палеонтологического института. Том 67. М.: Академия наук СССР, 1957
Agadjanian A., Kondrashov P. Gobiatherium (Dinocerata, Mammalia) from Kyrgyzstan: morphology and taxonomic position. Paleontological 33 (3), 1999
Rose K. D. The Beginning of the Age of Mammals. — Oxford: Johns Hopkins University Press, 2006