Казеивые — Caseidae
Класс †Синапсиды (Synapsida)
Подкласс †Пеликозавры (Pelycosauria)
Надотряд †Офиакообразные (Ophiacomorpha)
Отряд †Казеазавры (Caseasauria)
Семейство †Казеивые (Caseidae)
Казеивые (Caseidae Williston, 1911) — семейство амниот, входящие в состав подотряда казеазавров (Caseasauria Williston, 1911) согласно классической систематике Р. Рейса, либо отряда казеазавров (Caseasauria Williston, 1911) в альтернативной систематике М.Ф. Ивахненко. Известны из позднего карбона - средней перми Северной Америки и позднего карбона - средней (? поздней) перми Европы. Установлено порядка 17 родов.
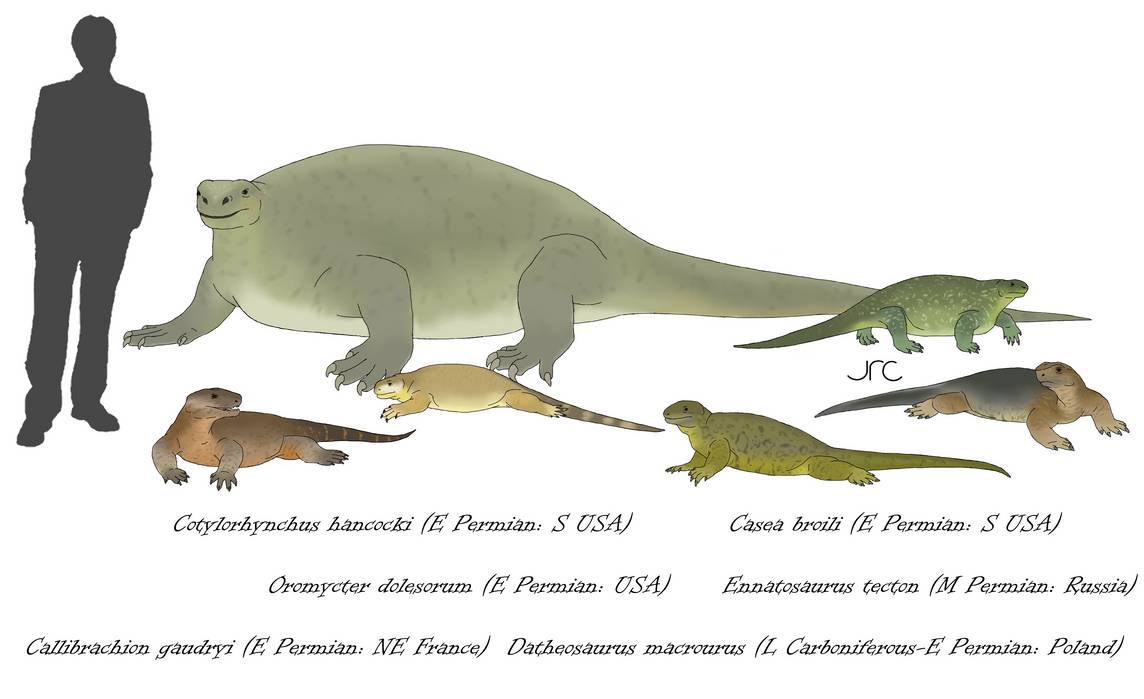
Разнообразие и относительные размеры казеивых (Caseidae). Автор — artbyjrc.
Общая длина составляет от 0,5 до 6 м, увеличиваясь на протяжении эволюционной истории.
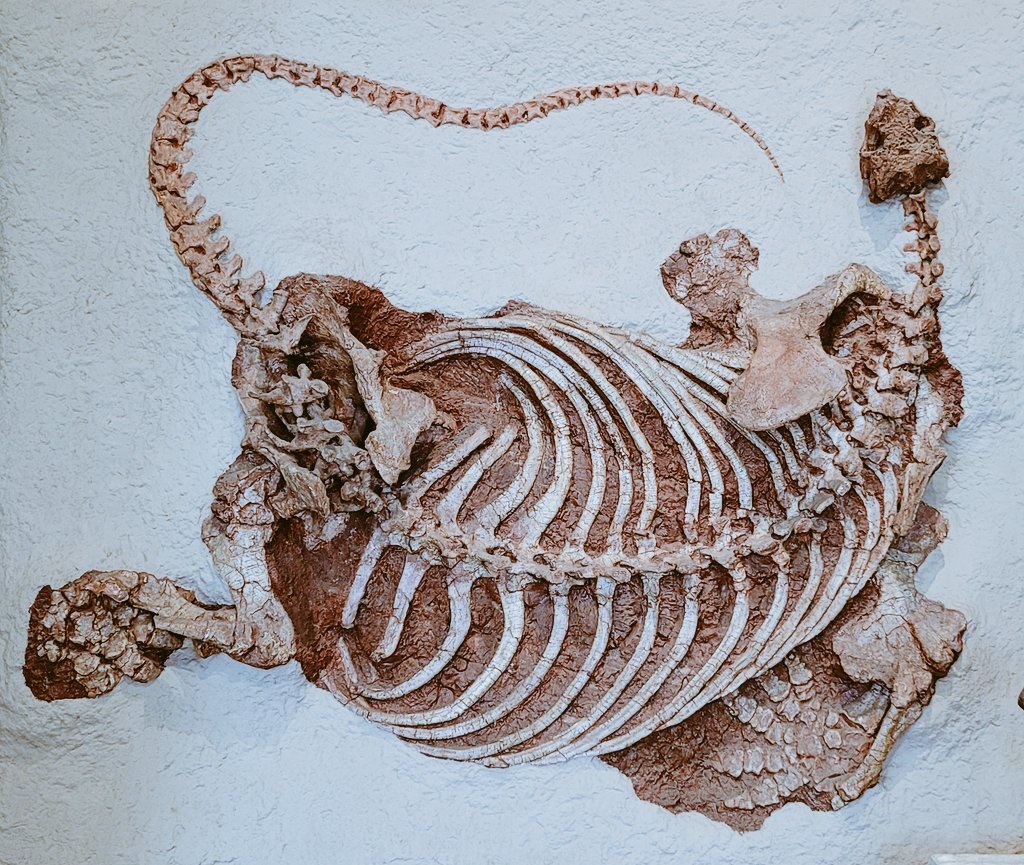
Составный скелет котилоринха (Cotylorhynchus romeri) (образец AMNH FARB 7517) из ранней перми штата Оклахома, США. Американский музей естествознания, Нью-Йорк.
Череп относительно размеров тела небольшой, а у поздних крупных представителей — очень маленький (при общей длине около 4 м — череп менее 20 см в длину). Череп короткий и низкий, расширяющийся к затылку. Наружные костные ноздри удлиненные и необычно крупные, иногда по размерам мало уступающие глазницам. Они занимают практически всю предглазничную часть черепа. Впрочем, такое увеличение ноздри в значительной мере кажущееся, поскольку большая ее часть перекрывается сильно увеличенной септомаксиллой, редко сохраняющейся на черепе. Септомаксилла выходит не только на задний, но и на верхний край ноздри, по которому широко сочленяется с носовой костью. Топографически септомаксилла имеет некоторое сходство с комплексом rostrale laterale и tectale superior кистеперых рыб. Обычно такой септомаксиллы у остальных известных в этом отношении тетрапод не бывает, хотя у многих пеликозавров она также хорошо развита (однако имеет при этом другое строение). В задней части ноздри могла находиться крупная латеральная носовая железа, для которой нельзя исключить и функцию солевыведения.

Череп с нижней челюстью котилоринха (Cotylorhynchus romeri) (образец OMNH 04329) из ранней перми штата Оклахома, США.
В отличие от большинства пеликозавров, слезная кость укороченная, что отчасти связано с укорочением морды и увеличением ноздрей. Ротовое отверстие смещено на нижнюю сторону черепа, так что верхняя челюсть заметно нависает над нижней. Височное окно крупное, ограниченное снизу контактом квадратно-скуловой и скуловой костей кости (как у Bolosaurida из капториноморф, или Lanthanosuchida из парарептилий). Формирование такого окна из "синапсидного" не доказано. Скуловой отросток чешуйчатой кости отсутствует. Верхняя затылочная кость небольшая. Пинеальное отверстие очень большое. Кости крыши черепа скульптированы россыпью небольших округлых ямок. Околозатылочные отростки чрезвычайно расширены. Челюстное сочленение расположено намного ниже уровня зубного ряда. На небе имеются многочисленные мелкие зубы, которые не концентрируются на специальных площадках и покрывают также парасфеноидный ростр. Нижняя челюсть меньше верхней, без засочленовного отростка. Ее симфиз образован как зубной, так и пластинчатой костями.
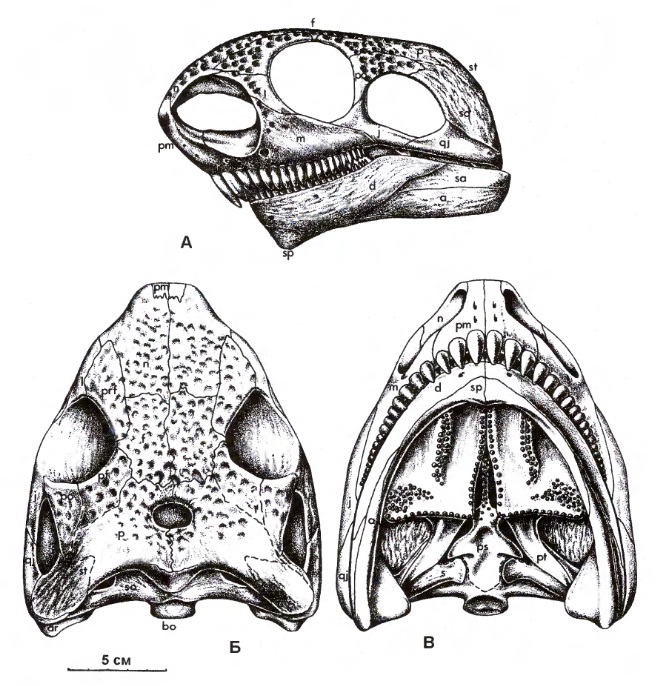
Прорисовки реконструированного черепа с нижней челюстью котилоринха (Cotylorhynchus romeri): сбоку (а), сверху (б) и снизу (в).
Зубная система также весьма специфична. По сравнению с эдафозаврами, число челюстных зубов уменьшено. Сами челюстные зубы однородные. Передние укрупненные, загнуто-конические, боковые и задние — более мелкие, продольно-уплощенные, у типичных представителей зазубренные по краям. При закрытии пасти верхние зубы без окклюзии перекрывают нижние. На венечной кости зубы обычно отсутствуют.
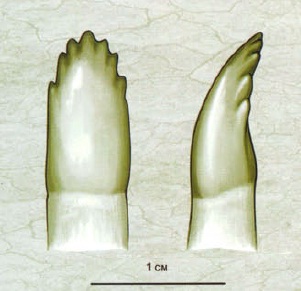
Прорисовки заднего зуба эннатозавра (Ennatosaurus tecton), спереди и сбоку.
Туловище широкое и приплюснутое, без спинного "паруса". Предкрестцовых позвонков 25, число крестцовых позвонков возрастает до 3-4. Тело атланта достигает уровня вентральной поверхности позвоночного столба. Шейные позвонки малы, туловищные умеренно удлиненные, катушкообразные, без вентрального киля. Интерцентры малы. Остистые отростки короткие. Поперечные отростки умеренно удлинены и смещены вперед и вверх по невральной дуге. Невральные дуги без вырезок над конечными отростками. Зигапофизы велики и заметно выступают латерально. Ребра сильно изогнуты, так что грудная клетка была весьма объемистой, бочкообразной. Поясничные ребра очень большие, сросшиеся с позвонками. Число крестцовых ребер внутри группы увеличивается с 3 до 4 пар. Реберный бугорок почти не развит. Хвост относительно длинный.

Смонтированный скелет котилоринха (Cotylorhynchus romeri) из ранней перми штата Оклахома, США.
Ключица дистально расширена. Лопатка короткая, дистально суженная. Супрагленоидное отверстие отсутствует. Подвздошная кость сильно разрастается вперед от вертлужной впадины. Конечности укороченные и мощные, их кости массивные. Проксимальные отделы конечностей умеренно удлиненны. Плечевая кость без эктэпикондилярного отверстия. Как и у эдафозавров, супинаторный отросток разросся дистально, полностью данное отверстие охватывая. Бедренная кость с Y-образным аддукторным гребнем. Кисть и стопа короткие и широкие (черта, постепенно усиливающаяся внутри группы). Особенно сильно расширен V-й палец стопы. Число фаланг всегда меньше, чем у других пеликозавров, и может сокращаться до 2 - 2 - 3 - 3 - 2. Когтевые фаланги большие.
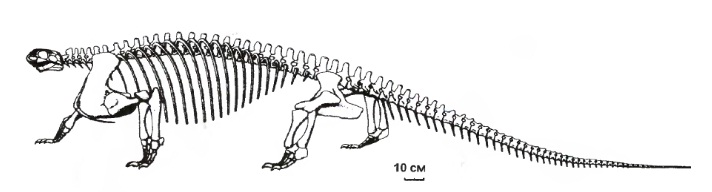
Прорисовка реконструированного скелета котилоринха (Cotylorhynchus romeri).
Образ жизни казеид не вполне ясен. Они не встречаются в обычных раннепермских фаунах США и, возможно, жили в особых местообитаниях. В последнее время казеиды обычно рассматриваются как факультативно-водные обитатели побережий мелких солоноводных бассейнов. Крупные виды могли быть почти полностью водными. Об этом косвенно свидетельствует губчатая структура их костной ткани, существенно облегчающая скелет. Новый смысл в данном свете обретает массивное туловище с короткой шеей и веслоподобными конечностями, которые могли использоваться казеидами для плавания. Объяснение находит и предположительно имевшиеся в ноздрях солевыводящие железы, напоминавшие таковые современных морских игуан Amblyrhynchus. Не исключается также, что для интенсивного дыхания во время активного плавания и погружений указеид была развита диафрагма. С другой стороны, многие их представители имели крупные когти и могли являться частично роющими.

Реконструкция внешнего облика котилоринха (Cotylorhynchus romeri). Автор — Г. Угуэто.
Ввиду отсутствия зубной окклюзии, эти животные могли лишь захватывать кормовые объекты или отрывать куски пищи, а затем проглатывать их практически не обрабатывая в ротовой полости. Судя по строению челюстных зубов, позднекарбоновый казеид Eocasea являлся насекомоядным, а у раннепермского Martensius рацион на протяжении жизни менялся от насекомоядного до всеядного. Характер питания более поздних, типичных представителей группы остается дискуссионным. В прошлом высказывалось предположение о питании казеид моллюсками. Долгое время популярным было мнение, что кормом им служили мягкие корни и разлагающиеся стволы отмерших растений. На это могут указывать огромные когтистые лапы, которыми было удобно выкапывать корневища и ломать вегетативные части растений. А грудная клетка расширена настолько, что обеспечивает достаточно пространства объемистому кишечнику, занятому ферментацией растительности.

Котилоринх (Cotylorhynchus romeri) в естественном окружении. Автор — М. Синица.
Мощный височный отдел предполагает сильный укус челюстей, которые двигались лишь в одной плоскости — вверх и вниз. Лепестковидные, с зазубренными краями челюстные зубы типичных казеид в целом сходны с зубами парейазавров, платеозавров и современных ящериц, питающихся мягкими частями растений. Боковые зубы челюстей несут не режущую мелкую зазубренность, а удлиненные зубцы — выступы, не несущие следов стирания, как это наблюдается и у парейазавров, вследствие чего для казеид так же выдвигается предположение о питании мягкой водной растительностью — возможно, водорослево-бактериальными матами. Отличием зубной системы казеид от таковой парейазавров является наличие у первых мощных долотовидных передних зубов, что, возможно, связано с захватом и отрыванием растительности. Мелкие и острые небные зубы помогали удерживать пищу во рту.

Котилоринх (Cotylorhynchus romeri) вырывает подземные части растений. Автор — М. Целески.
Водорослево-бактериальные маты нарастали на поверхности водоемов подобно ряске. Они прорастали грибами и мхами и образовывали густые прибрежные ковры и плавающие острова. В их толще сновало множество рачков, насекомых и их личинок. По сути, это был сверхпитательный растительно-мясной салат, для потребления которого годилась самая примитивная стратегия. Своей нижней челюстью, как ковшом экскаватора, казеид захватывал в рот комок мата, с помощью языка запихивал его поглубже в пасть, причем правильное направление движению задавали изогнутые назад небные зубы. Затем пасть захлопывалась, а кусок крепко зажимался между верхней и гораздо меньшей нижней челюстью. Обрезать кусок мата казеид не мог: его зубы были довольно тупыми. Перепилить тоже — он не умел жевать. Животное было способно лишь оторвать кусок. Затем оно мотало головой в разные стороны, пока все растительные волокна не разрывались. Так или иначе, казеиды стали одними из первых широко распространенных растительноядных наземных позвоночных.

Котилоринхи (Cotylorhynchus romeri) в естественном окружении. Автор — А. Атучин.
А. Ромер и Л. Прайс (1940 г.) поместили эдафозавров и казеид в один подотряд. Этим группам присущ ряд общих производных признаков, большинство которых можно связать с крупным размером тела, но специализация зубов к растительноядности у них совершенно различна. Если у них и был общий предок, то на уровне примитивного хищного пеликозавра. Р. Рейс (1981 г.) указывал, что эотириды и казеиды уникальны среди пеликозавров вытянутой в переднезаднем направлении наружной ноздрей, заостренным ростром и участием верхнечелюстной кости в формировании вентрального края глазницы. У обеих групп также короткая морда и низкий пластиновидный затылок, хотя эти признаки для них и не уникальны. Как указывал Э. Олсон (1968 г.), эотириды далеки от казеид природой своей зубной системы.
Семейство имеет следующий состав:
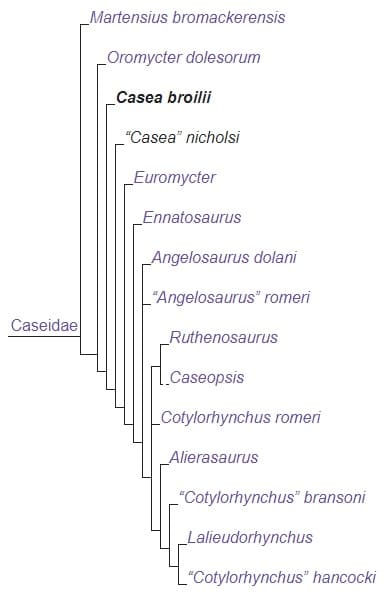
Кладограмма, иллюстрирующая родственные связи казеивых (по Вернебургу с соавт., 2022 г.).
Семейство имеет следующий состав:
род †Эоказеи (Eocasea Reisz et Fröbisch, 2014);
род †Мартенсии (Martensius Berman, Maddin, Henrici, Sumida, Scott et Reisz, 2020);
род †Датеозавры (Datheosaurus Schröeder, 1905);
род †Каллибрахионы (Callibrachion Boule et Glangeau, 1893);
род †Аризиерпетоны (Arisierpeton Reisz, 2019);
род †Оромиктеры (Oromycter Reisz, 2005);
род †Трихазавры (Trichasaurus Williston, 1913);
род †Казеи (Casea Williston, 1910);
род †Ангелозавры (Angelosaurus Olson et Beerbower, 1953);
род †Казеопсисы (Caseopsis Olson, 1962);
род †Казеоиды (Caseoides Olson et Beerbower, 1953);
род †Котилоринхи (Cotylorhynchus Stovall, 1937);
род †Евромиктеры (Euromycter Reisz, Maddin, Fröbisch et Falconnet, 2011);
род †Рутенозавры (Ruthenosaurus Reisz, Maddin, Fröbisch et Falconnet, 2011);
род †Алиеразавры (Alierasaurus Romano et Nicosia, 2014);
род †Лалиэвдоринхи (Lalieudorhynchus Werneburg, Spindler, Falconnet, Steyer, Vianey-Liaud et Schneider, 2022);
род †Эннатозавры (Ennatosaurus Efremov, 1956).
Ссылки
Казеиды (Википедия) перейти
Caseidae (Paleobiology Database) перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Иванов А. О., Черепанов Г. О. Ископаемые высшие позвоночные. — СПб.: Издательство Санкт-Петербургского университета, 2001
Ивахненко М. Ф. Тетраподы Восточно-Европейского плаката — позднепалеозойского территориально-природного комплекса. — Пермь, 2001
Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1 / Отв. ред. Ивахненко М. Ф. и Курочкин Е. Н. — М.: ГЕОС, 2008
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Нелихов А. Е. Ящеры Пинеги. — М.: Нужненко Д. Б., 2011
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГЕОС, 2009