Подкласс †Пеликозавры (Pelycosauria)
Надотряд †Сфенакообразные (Sphenacomorpha)
Отряд †Сфенакодонты (Sphenacodontia)
Клада †Гаптодонтиобразные (Haptodontiformes)
Семейство incertae sedis
Род †Кеномагнаты (Kenomagnathus)

В 1931 г. Норман Ньюэлл обнаружил окаменелость в Гарнеттском карьере. Примерно в 1932 г. Генри Лейн и Клод Хиббард собрали в этом местонахождении множество животных и растительных остатков. Среди них были скелеты примитивной диапсидной рептилии Petrolacosaurus kansensis, описанные в 1952 г. Фрэнком Пибоди. В надежде найти больше материала, полевая команда из Музея естественной истории Университета Канзаса провела раскопки в 1953 и 1954 гг. Ими были найдены отпечатки следов, остатки целакантовых рыб, несколько новых скелетов петролакозавра и окаменелости пеликозавров 3 разных родов. Один из частичных скелетов явно отличался от других. Пибоди сообщил об этих открытиях в 1957 г., заметив, что упомянутый скелет принадлежит примитивному сфенакодонтиду, но описания его не дал.
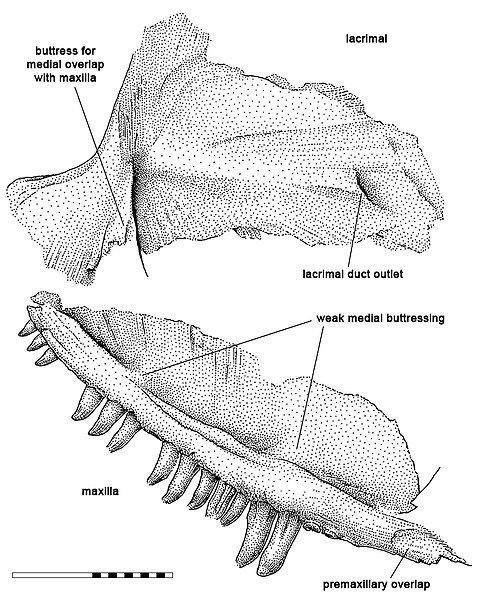
В 1965 г. Роберт Кэрролл обнаружил в карьере Гарнетт еще один сочлененный частичный скелет. Филип Карри приписал его примитивному сфенакодонтиду Пибоди, которого он в 1977 г. поместил в род Haptodus, установив для него новый вид Haptodus garnettensis. Все известные на то время экземляры Haptodus garnettensis были либо сильно раздроблены, либо являлись молодыми особями. В течение 1980-х гг. в карьере было обнаружено еще несколько экземпляров, включая взрослых и находящийся на пороге зрелости. Это позволило Мишелю Лаурин выделить отличительные признаки Haptodus garnettensis и включить их в филогенетический анализ, который показал, что данный таксон находится за пределами Sphenacodontidae. Лаурин опубликовал эти результаты в 1993 г. Среди дополнительных образцов имелся частичный череп, состоящий из левой верхнечелюстной и слезной костей, который был занесен в каталог Королевского музея Онтарио как ROM 43608.
Анализ приписываемых Haptodus garnettensis образцов, проведенный позже Фредериком Шпиндлером и его коллегами, показал, что они на самом деле принадлежат не одному, а от 4 до 6 самостоятельным таксонам, которые отличаются друг от друга главным образом строением челюстей и зубов. Исследователи также признали различия между "Haptodus" garnettensis и типовым видом Haptodus baylei. В своем выступлении на конференции 2013 г. Фредерик Шпиндлер, Кирстин Бринк и Грасиела Пиньейро предположили, что морфологические различия новооткрытых животных основаны на диетических предпочтениях, что делает их доисторическим аналогом Дарвиновых вьюрков.
В 2020 г. Шпиндлер официально поместил образец ROM 43608 в новый таксон, названный им Kenomagnathus scottae. Родовое наименование образовано греческими словами kenoma — "промежуток, пустота" и gnathos — "челюсть", что связано с наличием выразительной диастемы в верхнем зубном ряду. Видовой эпитет присвоен в честь препаратора Дайан Скотт, помогавшей Шпиндлеру в работе с окаменелостями.
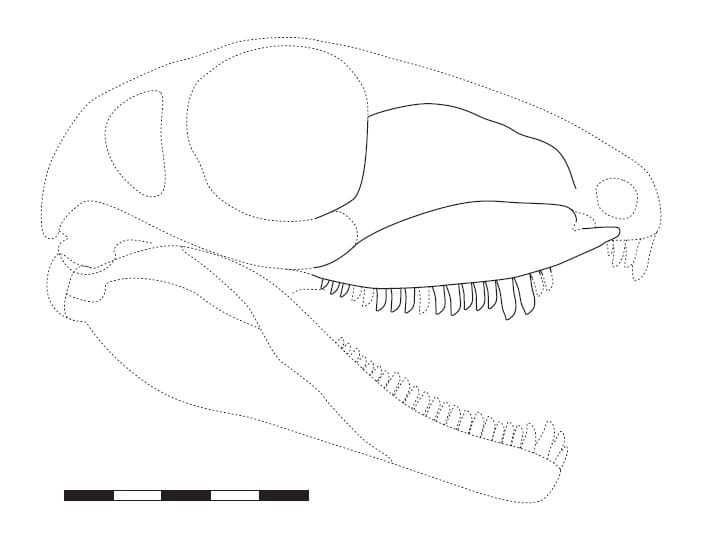
Прорисовка частично реконструированного черепа с нижней челюстью кеномогата (Kenomagnathus scottae) (по типовому образцу ROM 43608). Длина масштабной линейки — 5 см.
Kenomagnathus характеризуется высокой мордой, судя по конфигурации его верхнечелюстной и, особенно, слезной кости. Значительная высота слезной кости, окаймлявшей переднюю часть глазницы, также указывает на большой размер глаз. Нижний край верхнечелюстной кости у Kenomagnathus более выпуклый, чем у "Haptodus" garnettensis, и лишен вогнутой области (т. н. предклыковой ступени). Другой отличительной чертой является наличие диастемы — беззубого промежутка, охватывающего ширину по меньшей мере 3 зубов в передней части верхнечелюстной кости, где она истончена и не содержит зубные альвеолы. За диастемой расположены 2 предклыковых зуба, 2 больших клыка и по меньшей мере 14 заклыковых зубов (из которых на имеющемся образце сохранились 11). Также на верхнечелюстной кости имеются 2 слабо утолщенные области, или контрфорса, — одна над клыками, а другая чуть позади них. Из-за укороченной верхнечелюстной кости клыки вынесены вперед в большей степени, чем у родственных таксонов. Подобно "Haptodus" garnettensis, зубы высокие и почти прямые с бороздками на внутренней поверхности, достигающими кончиков, но у Kenomagnathus они более тонкие и притупленные на концах.
В своей диссертации за 2015 г. Шпиндлер привел предварительный филогенетический анализ "хаптодонтин", основанный на новом наборе характеристик, которые были собраны этим автором для определения их родственных взаимоотношений. После рассмотрения образцов, связанных с "Haptodus" garnettensis в противопоставление установленным в 2020 г. Kenomagnathus scottae и Tenuacaptor reiszi, Шпиндлер не мог решить, какой таксон занимает более базальное положение (т. е. является менее специализированным) — Kenomagnathus или Ianthodon schultzei. Однако выделение Tenuacaptor указало на более производное (т. е. более специализированное) положение внутри Sphenacodontia как Kenomagnathus, так и "Haptodus" garnettensis.
В 2020 г. Шпиндлер привел 3 признака, позволяющих отнести Kenomagnathus к Sphenacodontia: выпуклый нижний край верхнечелюстной кости, значительная высота слезной кости и притупленные зубы. Однако фрагментарность имеющегося материала усложняет определение взаимоотношений, так что Kenomagnathus вполне может располагаться за пределами группы Sphenacodontoidea. Это усугубляет то обстоятельство, что многие "хаптодонтины" очень похожи друг на друга, за исключением различий в черепных пропорциях и строении зубов.

Часть левого верхнего зубного ряда кеномогата (Kenomagnathus scottae) (типовой образец ROM 43608) с предклыковой диастемой.
Современные млекопитающие (за исключением китообразных) обычно демонстрируют гетеродонтность, т. е. обладают зубами нескольких различных типов. Среди их вымерших родственников наблюдается переход от животных с гомодонтными (однородными) зубами к животным с гетеродонтными зубами, связанный с развитием отчетливых "зон" вдоль зубного ряда. Клыковидные зубы — в частности, двойные клыки, такие как у Kenomagnathus, — довольно широко распространены среди базальных синапсид, а также других ранних амниот, наряду с увеличением первого зуба на предчелюстной кости в передней части черепа. Существует морфологический разрыв между базальными синапсидами, такими как "Haptodus" garnettensis, обладающими дополнительными предклыковыми зубами, но менее вариабельным зубным набором, и более близкими к млекопитающим терапсидами, у которых отсутствуют предклыковые зубы, а есть единственный набор клыков и гетеродонтность выражена сильнее. У терапсид предклыковые зубы часто заменены широкой диастемой.
У Kenomagnathus имеются как предклыковые зубы, так и диастема, что до некоторой степени заполняет промежуток между базальными синапсидами и терапсидами. Это может считаться иллюстрацией определенного этапа эволюционного перехода, который проходили обладающие диастемами синапсиды. Шпиндлер предположил, что диастема первоначально развилась в результате того, что верхнечелюстная и предчелюстная кости наклонялись друг к другу на предклыковой ступени, предотвращая удлинение предчелюстных зубов; это рассматривается как т. н. начальная диастема у "Haptodus" garnettensis, Ianthasaurus и Ianthodon. Такое строение передней части верхней челюсти могло позволить противостоящим нижним зубам вырасти длиннее и заполнить имеющийся промежуток. Однако Шпиндлер также отметил, что взаимосвязь между развитием предклыковой ступени, потерей зубов и расширением диастемы остается неясной, и что стиль питания этих животных также мог оказывать свое влияние.
Шпиндлер сравнил Kenomagnathus с загадочным синапсидом Tetraceratops, поскольку для обоих характерны короткая морда, большие глазницы и диастема в верхнем зубном ряду. Хотя это подразумевает, что черепа этих животных демонстрируют специализацию к определенному типу питания, Шпиндлер указывает, что их дальнейшие сравнение охватывает более широкий спектр возможностей. В частности, Tetraceratops явно более специализирован: его верхнечелюстная диастема не совпадает с положением нижних клыковидных зубов, а расположена немного позади них, и первый предчелюстной зуб выступал из передней части челюстей, когда они были сомкнуты. Шпиндлер высказал предположение, что Tetraceratops был дурофагом (т. е. питался беспозвоночными с твердым панцирем или экзоскелетом), основываясь в основном на строении его зубов. Ученый предположил аналогичный образ жизни и для Kenomagnathus, исходя из большой высоты его черепа, и допустил, что верхнечелюстную диастему могла покрывать жесткая ткань, на которую потенциально давил увеличенный нижний зуб при раздавливании кормового объекта.
Kenomagnathus входит в гарнеттский фаунистический комплекс позвоночных. Породы, в которых обнаружены его остатки, относятся к слою Рок-Лейк формации Стэнтон. Основание данного слоя состоит из углеродистых известняковых аргиллитов, умеренно слоистых и имеющих темно-серовато-коричневый цвет. Ископаемая флора этих породах представлена хвойными деревьями, кордаитами, семенными папоротниками, саговниками, хвощами и папоротниками. Выше расположен слой из раковин морских двустворчатых моллюсков толщиной 2-5 см, которые, вероятно, были отложены во время прилива. В совокупности это означает, что Kenomagnathus обитал на прибрежной равнине в хвойном лесу. Последующие слои горных пород становятся более неравномерными, а остатки наземных животных и растений в них все чаще заменяются окаменелостями морских беспозвоночных, что означает, что эстуарные воды постепенно затопляли регион по мере повышения уровня моря. Шпиндлер полагает, что Kenomagnathus мог питаться двустворчатыми моллюсками.