Приветствую Вас Гость | RSS
Век млекопитающих - Age of Mammals
Меню сайтаФорма входа |
Образ жизни По мнению Т. Рингстрема (1924 г.), особенности строения черепа и нижней челюсти с выступающими вперед бивнеобразными резцами, гипсодонтность коренных зубов, тенденция к развитию однопалости, а также массовые захоронения хилотериев в Восточной Европе и Азии свидетельствуют о их приуроченности к степным участкам с твердой почвой. Помимо прочего, данная точка зрения хорошо согласуется с изменением климата и распространением степей в позднемиоценовое время. Однако дальнейшее изучение показало большую приверженность хилотериев к влажным прибрежным и болотистым биотопам. О их питании болотной растительностью свидетельствует совкообразно расширенная нижняя челюсть, а также форма и расположение нижних резцов. В свою очередь, увеличение гипсодонтности щечных зубов, а также усложнение их жевательных поверхностей с изогнутыми эмалевыми петлями и высоким содержанием цемента может объясняться не питанием степными травами с большим количеством частиц кремнезема, а перетиранием болотной растительности со значительной примесью песка и земли. Тем не менее, заключение Рингстрема о тяготении хилотериев к степным ландшафтам может быть частично справедливо в отношении поздних представителей рода. На потребление жесткой растительности прямо указывает характер износа щечных зубов, жевательные поверхности которых являются уплощенными, а не корытообразными, как у фитофагов, специализирующихся на мягкой пище. Таким образом, характерная структура зубов хилотериев препятствовала чрезмерному их истиранию.  Хилотерии (Chilotherium orlovi) в естественном окружении. Автор — К.К. Флеров. Укорочение конечностей и, как следствие, низкое положение головы и туловища хилотериев, вероятно, было обусловлено их приспособлением к питанию травянистой растительностью и позволяло легче до нее дотягиваться. В отличие от более прогрессивного подсемейства Rhinocerotinae, к которому относятся и ныне живущие носороги, в процессе специализации у хилотериев не вытянулся затылок, что приводит к очень низкому положению головы, как у белого носорога (Ceratotherium simum), а сохранилось более горизонтальное положение черепа. Таким образом, скорее всего хилотерии были обитателями хорошо увлажненных речных долин и приозерных равнин, где питались прибрежной и водной растительностью. Очевидно, расширенная нижняя челюсть с бивнеобразными резцами была приспособлена к выкапыванию корневищ и луковиц околоводных растений из болотистого грунта. У хилотериев, вероятно, также имелись мясистые губы, служившие для захвата густых трав и водной растительности. Судя по всему, по образу жизни эти носороги напоминали современного гиппопотама (Hippopotamus amphibius), были хорошо приспособлены к хождению по вязкой почве и, вероятно, неплохо плавали. Необходимо отметить, что степень привязанности разных видов к прибрежным биотопам была различной. Значительные скопления ископаемых остатков может свидетельствовать о стадности этих носорогов. Стадный образ жизни обеспечивал относительную безопасность отдельных особей и успешное выживание молодняка, находившегося под защитой старших.  Динокрокута (Dinocrocuta gigantea) охотится на хилотерия (Chilotherium wimani). Автор — Х. Ласерда. Мало что известно о естественных врагах хилотериев, но ввиду значительных размеров этих животных можно предположить, что, как и современные носороги, они нечасто подвергались нападениям плотоядных. Один из черепов Chilotherium wimani, происходящий из котловины Линься, несет зажившие повреждения, оставленные зубами какого-то крупного хищника — возможно, динокрокуты (Dinocrocuta gigantea). 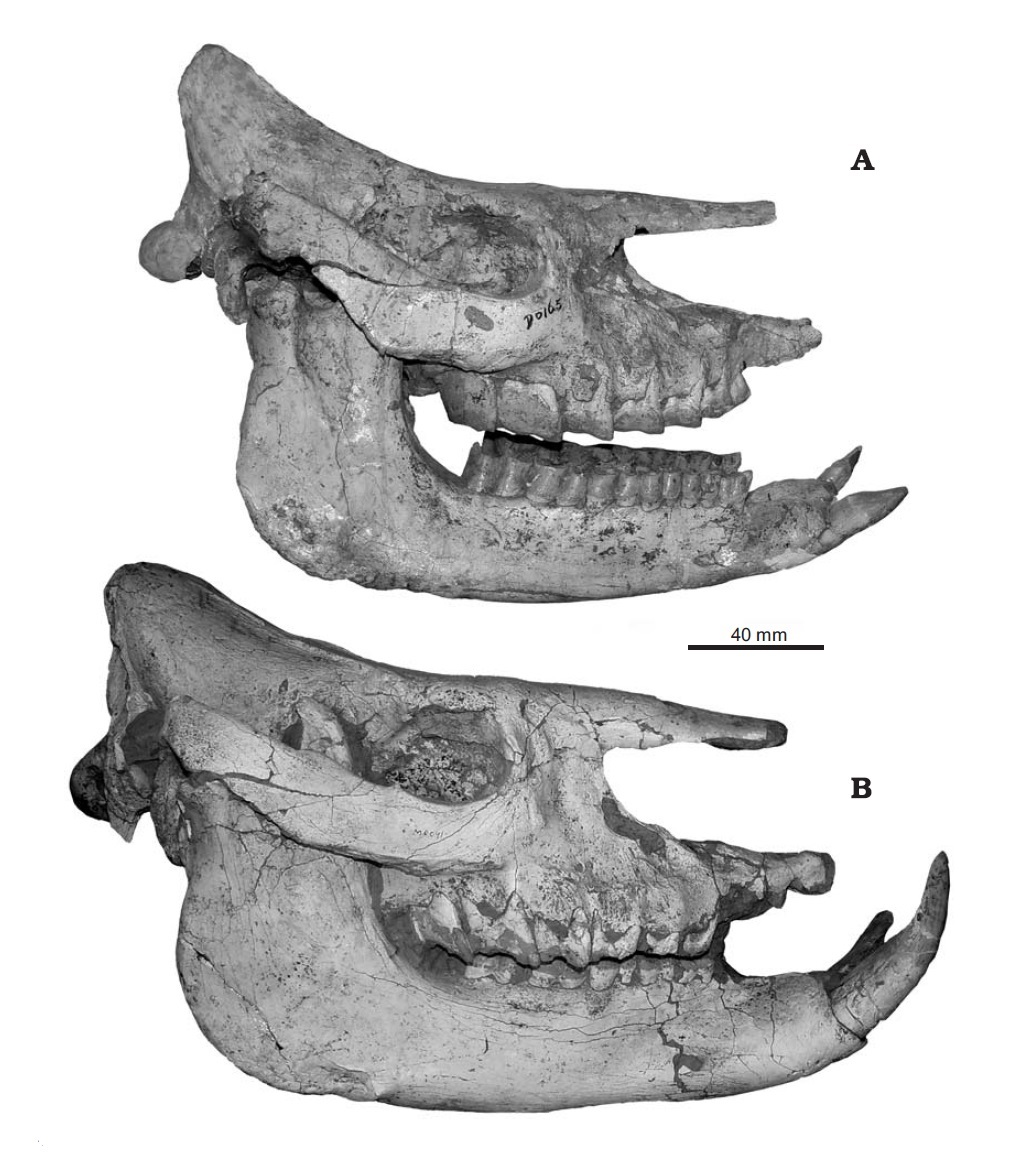 Черепа с нижними челюстями самки (А) и самца (В) хилотерия (Chilotherium wimani). Большие и острые наружные нижние резцы дополнительно могли использоваться для защиты от хищников и в качестве турнирного оружия. Поскольку у Chilotherium wimani зафиксирован половой диморфизм — в частности, бивни самцов длиннее и мощнее — вполне вероятно предположение, что между ними происходили стычки из-за самок, а также при выяснении иерархического статуса. Доказательством этого могут служить более изношенные, а иногда и отломанные резцы самцов Chilotherium wimani. Судя по всему, половой диморфизм был характерен и для других видов рассматриваемого рода. 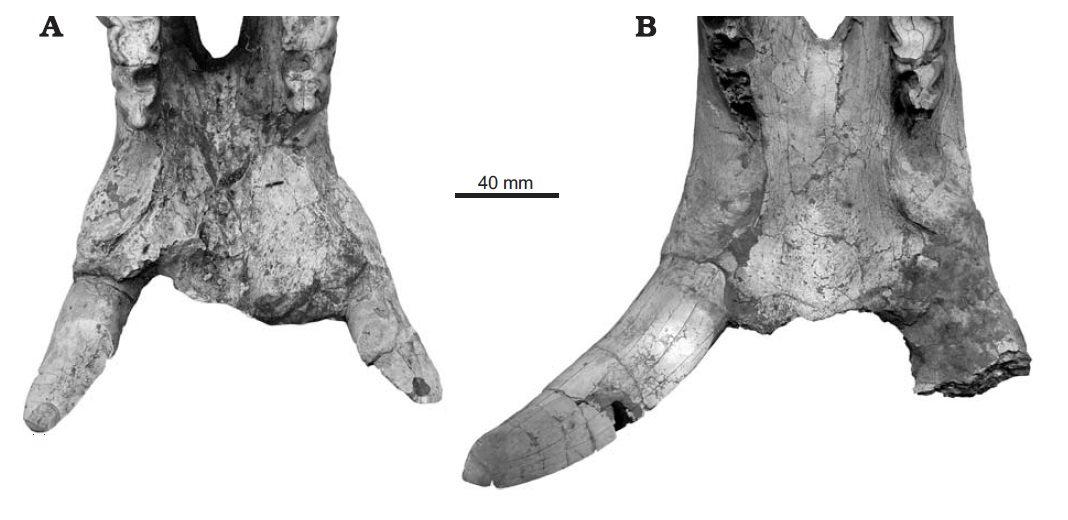 Сифизный отдел нижней челюсти самки (А) и самца (В) хилотерия (Chilotherium wimani). Численность хилотериев, судя по частоте встречаемости их остатков, была чрезвычайно высокой. Они преобладают в местонахождениях гиппарионовых фаун юга Украины, Казахстана и Китая. Возможно, кладбища этих животных образовались в засушливый период на берегах пересыхающих водоемов, а не в результате нагромождения остатков поверхностными водами в течение многих сезонов. Анализ остатков почти 200 хилотериев из отложений свиты Люшу в котловине Линься демонстрирует необычно высокий процент взрослых животных возрастом от 14 до 32 лет, которые составляют более 2/3 всех особей, в то время как молодые и старые животные редки. Поскольку все эти хилотерии, жившие на берегу озера, погибли по естественным причинам и ни в одном не стали жертвами природных катастроф, неясно, почему так много взрослых особей сохранилось в палеонтологической летописи.   Коллекции черепов с нижними челюстями Chiloterium из массовых захоронений Китая. Происхождение, эволюция и вымирание Длительное время род Chilotherium числилился в составе подсемейства Teleoceratinae (ныне — триба Teleoceratini), однако в дальнейшем выяснилось, что некоторые общие с последними морфологические признаки (в частности, укороченные конечности) развились у представителей эволюционной линии Chilotherium независимо в связи со сходным образом жизни. В результате ревизии Антуана и Сарача (2005 г.) род был отнесен к трибе Aceratheriini, а Дэн (2005 г.) выделил его в отдельную трибу Chilotheriini в составе подсемейства Aceratheriinae. 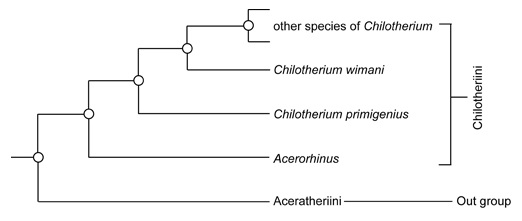 Один из вариантов филогении рода Chilotherium. Поскольку первые представители рода — наиболее ранний Chilotherium рrimigenius и живший несколько позднее Chilotherium wimani — обнаружены в Китае, центром происхождения этих носорогов можно считать Восточную Азию, откуда они в дальнейшем расселились до Балканского полуострова и Малой Азии на западе и до Юго-Восточной Азии на востоке. В северо-западном направлении непреодолимой естественной преградой на пути расселения этих коротконогих носорогов стали горы Карпаты. Вероятно, рассматриваемый род возник от более примитивного китайского рода Acerorhinus, ранее включаемого в род Chilotherium. Довольно близок к последнему также Subchilotherium, в прошлом также относимый к роду Chilotherium. Возможно, они представляют собой 2 параллельные линии, происходящие от общего предка. Данные китайских палеонтологов (Дэн, 2006 г. и др.) свидетельствуют о том, что предковой формой или близкой к таковой мог быть наиболее примитивный вид рода Chilotherium primigenius, давший начало Chilotherium wimani, от которого в свою очередь могли произойти Chilotherium xizangensis, Chilotherium persiae и Chilotherium kiliasi. Более поздней видовой группой рода являются Chilotherium anderssoni, Chilotherium habereri, Chilotherium samium, Chilotherium schlosseri и Chilotherium kowalevskii. Самым поздним и прогрессивным представителем рода был павлодарский Chilotherium orlovi (Байшашов, 2008 г.). Эволюция хилотериев протекала в неразрывной связи с влажными, прибрежными и заболоченными биотопами, которые населяли эти носороги. Род оказался весьма успешным, он существовал на протяжении по меньшей мере 8 млн. лет на значительной территории Евразии и дал веер местных разновидностей. Расцвет рода пришелся на конец миоцена. 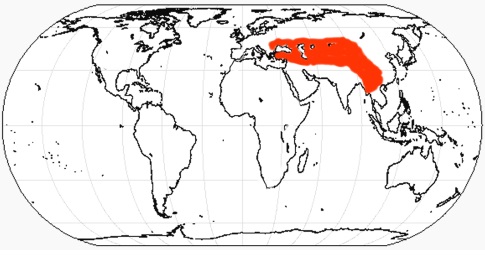 Ареал распространения ископаемых остатков рода Chilotherium. Хилотерии являются одним из важнейших компонентов гиппарионового фаунистического комплекса Восточной Палеарктики. Их остатки известны из многих местонахождений Греции, Болгарии, Молдавии, Румынии, юга Украины (в том числе и со дна Черного моря), юга европейской части России, Турции, Ирана, Северного и Восточного Казахстана, Киргизии, Монголии и Китая. В раннеплиоценовое время хилотерии повсеместно исчезают. Очевидно, это было связано с происходившим на протяжении плиоцена иссушением климата Евразии. Когда оно вызвало резкое сокращение приводных зарослей, эти болотные носороги, потеряв свой привычный мир, вымерли.  Смонтированные скелеты Chilotherium из массового захоронения в Фугу, провинция Шаньси, Китай. |
Поиск |