"Креодонты" — Creodonta
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Клада Скротиферы (Scrotifera)
Клада Феревунгуляты (Fereuungulata)
Клада Ферэ (Ferae)
Клада Всехищные (Pan-Carnivora)
"Креодонты" (Creodonta)
Отряд †Оксиенодонты (Oxyaenodonta)
Отряд †Гиенодонты (Hyaenodonta)
"Креодонты" (Creodonta Cope, 1875) — в прошлом отряд высших млекопитающих (Eutheria Gill, 1872), а в настоящее время парафилетическая группа клады всехищные (Pan-Carnivora Flynn, Wyss et Wolsan, 2020), происходящая из среднего палеоцена - позднего олигоцена Северной Америки, среднего палеоцена - позднего миоцена Азии, раннего эоцена - раннего миоцена Европы и позднего палеоцена - среднего миоцена Африки. Данная группа объединяет 2 отряда всехищных дохищникоподобного уровня — оксиенодонтов (Oxyaenodonta Van Valen, 1971) и гиенодонтов (Hyaenodonta Van Valen, 1967), ранее включаемых в отряд креодонтов в качестве семейств.
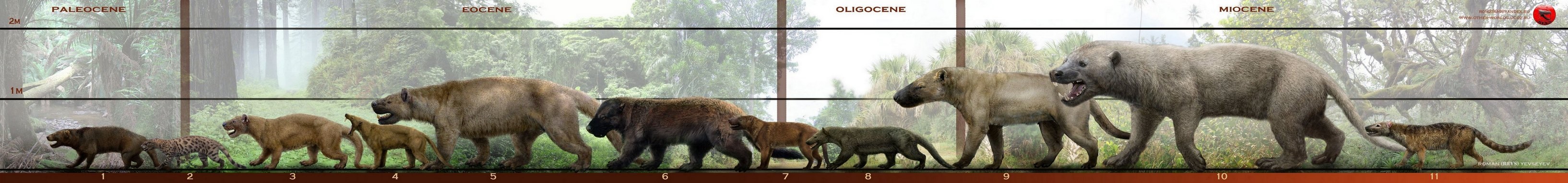
Разнообразие "креодонтов": 1 — палеониктис (Palaeonictis peloria) (Palaeonictinae, Oxyaenidae, Oxyaenodonta), поздний палеоцен Северной Америки; 2 — оксиена (Oxyaena lupina) (Oxyaeninae, Oxyaenidae, Oxyaenodonta), поздний палеоцен - ранний эоцен Северной Америки; 3 — патриофелис (Patriofelis ferox) (Oxyaeninae, Oxyaenidae, Oxyaenodonta), ранний эоцен Северной Америки; 4 — апатэлюр (Apataelurus kayi) (Machaeroidinae , Oxyaenidae, Oxyaenodonta), средний эоцен Северной Америки; 5 — саркастодон (Sarkastodon mongoliensis) (Oxyaeninae, Oxyaenidae, Oxyaenodonta), средний эоцен Азии; 6 — гемипсалодон (Hemipsalodon grangeri) (Hyainailourinae, Hyainailouridae, Hyaenodonta), поздний эоцен Северной Америки; 7 — гиенодон (Hyaenodon horridus) (Hyaenodontinae, Hyaenodontidae, Hyaenodonta), поздний эоцен - ранний олигоцен Северной Америки; 8 — птеродон (Pterodon dasyuroides) (Hyainailourinae, Hyainailouridae, Hyaenodonta), средний олигоцен Европы; 9 — гиенодон (Hyaenodon weilini) (Hyaenodontinae, Hyaenodontidae, Hyaenodonta), ранний миоцен Азии; 10 — мегистотерий (Megistotherium osteothlastes) (Hyainailourinae, Hyainailouridae, Hyaenodonta), ранний - средний миоцен Северной и Восточной Африки; 11 — диссопсал (Dissopsalis carnifex) (Teratodontinae, Hyaenodonta), средний - поздний миоцен Азии. Автор — Р. Евсеев.
Этимология
Название группы образовано древнегреческими словами κρέας — "мясо" и ὀδούς — "зуб", означая в переводе "мясозубы". Оно указывает на питание ее представителей сугубо животной пищей.
История изучения
Впервые ископаемые остатки "креодонтов" были обнаружены в 1820-е гг. в позднеэоценовых отложениях Парижского бассейна. Во 2-й половине XIX - начале XX вв. последовали многочисленные находки из стран Западной Европы и США, были описаны десятки новых родов и видов. Первые находки "креодонтов" в Азии были сделаны в миоценовых отложениях Сивалика и формации Нагри. Основную массу наиболее информативных находок удалось получить в ходе работ Центрально-Азиатской Американской экспедиции (1920-30 гг.).
Длительное время ранние находки черепных фрагментов из Франции, принадлежащих Hyaenodon и Pterodon, относили к сумчатым (Marsupialia) или к хищным (Carnivora). Мнение о принадлежности к Eutheria утвердилось благодаря работам Г. Фийоля (1874 г.). Э. Коп (1875 г.) предложил название Creodonta, присвоив им подотрядный ранг в составе отряда насекомоядных (Insectivora). М. Шлоссер (1887 г.) повысил ранг Creodonta до отрядного, включив в его состав, кроме креодонтов (Oxyaenidae, Proviverridae), еще 3 семейства (Arctocyonidae, Mesonychia, Amblictonidae). Вортман (1901 г.) выделил Creodonta в качестве подотряда в составе Carnivora. У. Мэтью (1902, 1909 гг.) понимал Creodonta более широко. В современном понимании объема таксона, Creodonta соответствовали выделенному им семейству Pseudocreodi.
В дальнейшем объем и ранг "креодонтов" неоднократно пересматривался (Осборн, 1910 г.; Грегори, 1939 г.; Симпсон, 1945 г.; Ромер, 1966 г. и др.). Большинство исследователей в разработке систематики плотоядных млекопитающих использовали в качестве диагностических признаки строения зубного аппарата. Однако, как относительно недавно было показано, они приобретались конвергентно, в процессе специализации челюстного аппарата. Привычный в конце XX - начале XXI вв. ранг и объем "креодонтам" придал М. Маккенна (1975 г.), выделив их как отряд в составе cyперотряда Creofaga. В настоящее время таксон Creodonta упразднен ввиду установления его парафилетичности. При этом Hyaenodonta считаются сестринской группой по отношению к кладе Carnivoramorpha, а вместе они образуют сестринскую кладу по отношению Oxyaenodonta.
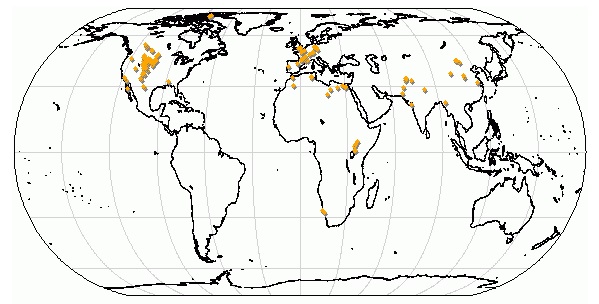
Карта основных местонахождений ископаемых остатков "креодонтов".
К сожалению, большинство "креодонтов" известны только по отдельным костям и фрагментам черепов. Их скелеты исключительно редки. Однако до нас все же дошли полные остатки американских оксиенодонтов (Oxyaena, Patriofelis) и гиенодонтов (Sinopa, Tritemnodon, Hyaenodon). Особенно многочисленны остатки Hyaenodon, что свидетельствует о этом роде как об одной из наиболее массовых форм палеогеновых хищников.
Систематика
В группу "креодонтов" условно объединяются 2 отряда — оксиенодонты (Oxyaenodonta) (порядка 15 родов) и гиенодонты (Hyaenodonta) (около 90 родов). Они отличаются друг от друга по строению зубочелюстного аппарата, стопы, голеностопного сустава и когтевых фаланг. Гиенодонты были более адаптивны и многочисленны, особенно в Старом Свете; как правило, представители этой группы имели собакоподобный или гиеноподобный облик и были лучше приспособлены к активному передвижению. Оксиенодонты, получившие распространение преимущественно в Северной Америке, в большинстве своем были относительно менее крупными и обладали удлиненным телом на коротких стопоходящих конечностях, напоминая по своему сложению енотов, медведей и кошек.
Уже у самых ранних оксиенодонтов зубная система была достаточно специализированной, тогда как архаичное плантиградное строение автоподия они сохранили на протяжении всей своей истории. У гиенодонтов, напротив, в отличие от оксиенодонтов, исходное строение зубной системы было примитивным, и в процессе эволюции происходили прогрессивные изменения как зубочелюстного, так и локомоторного аппаратов. Такое комплексное развитие обусловило большую экологическую пластичность гиенодонтов и позволило им успешно освоить новые территории и ландшафты.
Строение и функции
В ходе эволюции "креодонтов" реализовались различные стратегии достижения высокого уровня специализации и сформировались специализированные хищники всех размерных классов. Среди них встречались как карлики величиной с куницу, так и гиганты, способные соперничать своими размерами с самыми большими медведями (длина черепа — от 5 см у Lesmesodon до 67 см у Megistotherium). С течением времени представители обоих отрядов становились крупнее. Морфологическая организация "креодонтов" обладала рядом прогрессивных особенностей, но в общих чертах оставалась достаточно примитивной, сохраняя много архаических черт, сближающих их с насекомоядными мезозойскими предками.
Череп "креодонтов" большой относительно их тела, лицевой отдел как правило вытянут и почти равен по длине мозговому. Со временем черепа Oxyaenodonta приобретают ярко выраженный хищный облик — сильно укорачивается морда и раздвигаются в стороны скуловые дуги, у Hyeanodontidae, напротив, несмотря на резко хищный зубной аппарат, до конца сохраняют вытянутый узкий череп с относительно длинным и низким профилем. Особенно большими, почти непропорциональными головами обладали крупные представители группы, для которых длина головы соотносится с общей длиной тела в пропорции примерно 1 : 4. Для "креодонтов", как и для всех плотоядных млекопитающих, была характерна частичная редукция и специфическая дифференциация мышц предглазничного комплекса лицевой мускулатуры, очевидно, связанная со специализацией к захвату и удержанию крупной жертвы.
Примечательно расположение глазниц некоторых высокоспециализированных Hyaenodontidae, в частности представителей рода Hyaenodon. Как и у копытных, у них глаза находятся на боковых поверхностях черепа, что обеспечивает чрезвычайно широкий угол обзора. Очевидно, такая особенность была связана со сверххищным образом жизни этих "креодонтов" и как-то помогала им на охоте.
Морфология слуховой области наделена существенным сходством с Carnivora. Барабанная кость кольцевидная, костного слухового канала нет. Строение слухового пузыря архаично, частично он был построен из каменистой и чешуйчатой костей, однако при этом большая передне-латеральная часть оставалась хрящевой. Пока не известно, был ли второй центр окостенения слуховой капсулы, подобно Feliformia. У "креодонтов" имеется алисфеноидный канал, как у Caniformia. Заглазничное сужение значительное; верхние заглазничные отростки имеются почти всегда, нижние — отсутствуют; позади глазницы нет костной дуги.
Благодаря особенностям строения, челюстной сустав "креодонтов" позволял очень широко открывать пасть, но допускал только вертикальные движения нижней челюсти. Именно в связи с этим обстоятельством черепа большинства "креодонтов" обладают сильно развитым сагиттальным гребнем и относительно слабыми скуловыми дугами. Нижняя челюсть массивна, ее венечный отросток высокий. Параллельно плотоядной специализации щечных зубов в развитии челюстного аппарата "креодонтов" существовала тенденция к изгибу профиля щечного зубного ряда в верхне-нижнем направлении. Это обусловлено механическим выигрышем в разрезании пищевого объекта при поочередной окклюзии пар режущих зубов от P4/ml к М2/mЗ.

Череп с нижней челюстью гиенодона (Hyaenodon mustelinus).
Зубная система "креодонтов" очень своеобразна и по некоторым признакам конвергентно сходна с таковой Miacoidea, а также Borhyaenidae и Dasyuridae. Для нее присуща сильно выраженная мелкозубость (особенно по сравнению с современными хищниками). Зубная эмаль обладает специфической косой слоистостью, что делало ее более устойчивой. Судя по строению зубов, "креодонты" (в отличие от Didymoconida, Arctocyonia и Mesonychia) являлись высокоспециализированными хищниками.
Любопытно, что только у них наблюдается сокращение числа резцов. У ранних представителей отряда еще присутствует их полное число, они небольшие, мало различающиеся по величине и расположены в поперечном ряду. У высокоспециализированных форм, с ярко выраженным плотоядным типом коренных зубов, резцы претерпевают существенные изменения и достигают гораздо большей дифференциации, нежели у Carnivora. Из нижних i1 и i 3 уменьшаются в размерах, становятся впереди или позади i2, иногда исчезают. Из верхних редуцируются или исчезают I1 (у Pterodon) и I2, а I3 иногда сильно увеличивается и приобретает функцию дополнительного клыка (у Megistotherium), что свидетельствует о значительной хищнической специализации. У карнивор все иначе: резцов всегда по 3 пары сверху и снизу (единственное исключение — медведь-губач (Melursus ursinus), питающийся преимущественно муравьями).
Клыки "креодонтов" большие, заостренные, с одним корнем, зачастую массивные. Именно увеличенные клыки занимают освободившееся от резцов пространство. Получается, что эволюционный процесс "креодонтов" был направлен на снижение расстояния между клыками. Зачем — до конца непонятно. Возможно, это увеличивало силу укуса: сокращение челюстной мускулатуры более эффективно вгоняло клыки в плоть жертвы.
Предкоренные бугорчатые и относительно просто устроенные, задние верхние иногда с низким протоконом. Коренные зубы "креодонтов" изначально были построены по примитивному трибосфеническому типу, но в процессе эволюции они в большинстве случаев претерпели существенные изменения. Нижние коренные обладают высокими тригонидами и узкими талонидами; тригониды зачастую шире, нежели талониды; талониды нередко уменьшены, особенно на задних коренных. Нижний хищнический зуб обычно имеет уменьшенный метаконид и секториальный паракристид. Верхние коренные треугольной формы, с соединяющимися параконом и метаконом, небольшим конулем и хорошо заметным, но зачастую уменьшенным протоконом. Метастиль верхних хищнических зубов большой и соединенный с метаконом. Последний верхний коренной обычно уменьшен или полностью отсутствует, размеры последнего нижнего коренного варьируют от очень крупных до совсем небольших, вплоть до полной его редукции.
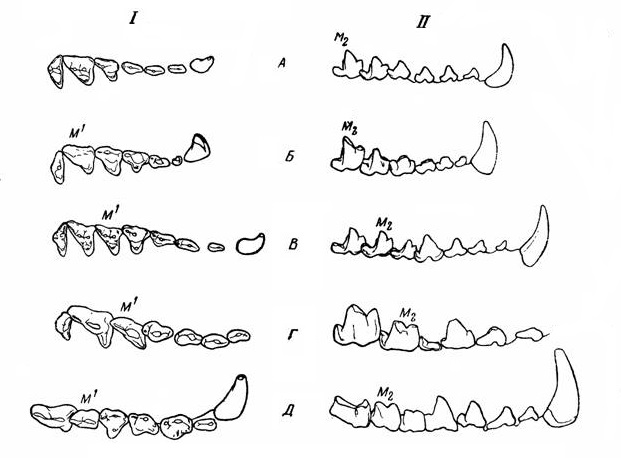
Схемы строения разных типов щечных зубов "креодонтов": I верхние, II— нижние; А — Thinocyon (Hyaenodonta), Б — Oxyaena (Oxyaenodonta), В — Sinopa (Hyaenodonta), Г — Pterodon (Hyaenodonta), Д — Hyaenodon (Hyaenodonta).
Для более эффективного разделывания животного корма щечной зубной ряд "креодонтов" эволюционировал в превосходный режущий аппарат, по действию напоминающий ножницы. Путем секториальной специализации некоторые зубы трансформировались в хищнические, обладающие острыми режущими поверхностями. Преобразованию примитивного бугорчатого зуба в режущий хищнический подверглись разные щечные зубы: у Oxyaenidae сильнее всего — М1 и m2, хотя на ранних стадиях их эволюции этим процессом частично захвачены также Р4 и m1. m1 иногда даже больше, чем m2 (подсемейство Palaeonictinae); эти зубы в дальнейшем снова упрощаются; М3, а позже и М2, так же как и m3, исчезают. У Hyaenodontidae трансформируются М1-2 и m2-3; все эти зубы на высших стадиях имеют режущий характер, но сильнее всего он выражен большей частью на М2 и m3, которые у поздних форм к тому же делаются более крупными, чем предшествующие; М3 уменьшается и исчезает. В обоих случаях наиболее специализирована задняя пара коренных зубов.
Специализация хищнических зубов привела у "креодонтов" к редукции давящих и перетирающих поверхностей, сохранившихся у настоящих хищных. Кроме того, у карнивор хищническими являются Р4 и m1, что кажется эволюционно более выгодным. Близость хищнических зубов "креодонтов" к заднему концу челюстей препятствовала такому значительному увеличению их в длину, как у наиболее хищных представителей хищных — кошек и гиен. Однако наиболее важным для дальнейшей эволюции "креодонтов" оказалось то, что их далеко расположенные хищнические зубы сделали затруднительным потребление какой-либо пищи, помимо мясной. Таким образом, в силу наличных морфологических предпосылок "креодонты" могли специализироваться исключительно в указанном направлении, в то время как зубной аппарат карнивор изначально является более многофункциональным и в ряде случаев позволяет разнообразить рацион растительным кормом.
Туловище "креодонтов" было приземистым и удлиненным, а конечности относительно короткими и мощными, практически всегда пятипалыми. Ключица как правило хорошо развита. Плечевая кость в большинстве случаев с внутренним, а иногда и со средним надмыщелковыми отверстиями; локтевая свободная, немногим тоньше лучевой, лучевая — с нерасчлененной верхней поверхностью. Ладьевидная, полулунная и центральная кости запястья почти всегда раздельны. Бедренная кость большей частью с III-м вертелом. Астрагал имеет заднее отверстие; его блок с неглубокой бороздой; имеется ясная косо направленная шейка, выпуклая головка сочленяется только с ладьевидной костью заплюсны, изредка также и с кубовидной. Ладьевидная и кубовидная кости заплюсны раздельны.
Стопа "креодонтов" (особенно у Oxyaenodonta) устроена крайне архаично. Oxyaenodonta и ранние Hyaenodonta (Proviverrinae, Limnocyoninae) изначально были стопоходящими и обладали достаточно широко расставленными плюсневыми костями и пальцами. У более поздних форм (Hyainailouridae, Hyaenodontidae) параллельно с некоторым удлинением конечностей пальцы собрираются "в комок", как и у большинства наземных хищников, приспособленных к бегу. У "креодонтов" наблюдается уклон в непарнопалость — III-й палец длиннее других. Последние фаланги несколько сжаты с боков (преимущественно у Hyaenodontidae) или расширены и расщепленные на конце сагиттальной щелью (чаще у Oxyaenodonta), когти без резкого заострения на конце. У Oxyaenodonta когтевые фаланги плоские, а когти были практически копытцеобразными. У Hyaenodontidae когти являлись маленькими и узкими, словно собачьи, только еще мельче. В общем когти "креодонтов" были невтяжными, тупыми и не годились для захвата жертвы.

Реконструированные правые передние лапы "креодонтов": слева — гиенодонт оксиенодон (Oxyaenodon dysodus), справа — оксиенодонт патриофелис (Patriofelis ferox). Автор — Р. Евсеев.
Небольшая толстостенная мозговая полость содержала соответствующий ее размерам примитивный головной мозг. Длительно время он считался сравнительно маленьким, но позже было убедительно продемонстрировано, что относительные размеры головного мозга "креодонтов" увеличивались так же быстро, как и у ранних хищных, параллельно расширению его новой коры (неокортекса). От самых первых "креодонтов" до более поздних представителей группы довольно четко прослеживается постепенное усложнение структур головного мозга. Однако в целом у "креодонтов" сохранялись почти гладкие и относительно крупные полушария большого мозга, не покрывающие мозжечка и крупных обонятельных луковиц, у многих были хорошо развиты височная и грушевидная доля, отвечающая за обоняние. Однако почти не вызывает сомнений, что даже при относительно гладком мозге у этих животных были неплохо развиты ассоциативные зоны, обеспечивавшие достаточно высокий уровень интеллекта, необходимый при их ярко выраженном хищном образе жизни. Мозжечек отделялся от остального мозга тенториумом.
Исследования эндокранов позволили установить, что некоторые "креодонты" характеризовались хорошо развитыми органами чувств (особенно зрением и обонянием, что проявляется в существенном увеличении соответствующих долей мозга и отделов новой коры) и обладали совершенной моторикой движений. Топологическая организация борозд и извилин неокортекса "креодонтов" относится к генерализованному продольному типу — они ориентированы параллельно сагиттальной оси (архаичная черта, присущая также ранним псовым). Специализация неокортекса сопровождалась морфологическими изменениями, характерными для всего отряда с сохранением продольной ориентации борозд и извилин. Отличия исследованных таксонов в специализации неокортеса кореллируют с различиями в строении челюстного аппарата. Гиперспециализация нервной системы, а не челюстного аппарата, сделала часть "креодонтов" уязвимой для изменений условий обитания и добывания пищи. Формирование адаптивных зон, связанное с глубокими изменениями в организации поведения и строении неокортекса "креодонтов", привело к сужению возможностей представителей отряда, что, видимо, и явилось одной из причин его вымирания.
Эволюционная история
Общими предками "креодонтов" (Creodonta) и настоящих хищных (Carnivora) являются пока неустановленные эвтерии, жившие в самом конце мела или начале палеоцена. Из предполагаемых синапоморфий обеих групп можно упомянуть наличие костного тенториума (перегородки, отделяющей мозжечок) и некоторых общих черт в строении основания черепа и предплюсны; указывалось также на присутствие синапоморфий в строении посткраниального скелета "креодонтов" и базальных хищноподобных типа Viverravus и Miacis. Все же, убедительно объяснить наличие общих морфологических признаков у "креодонтов" и карнивор посредством синапоморфий пока не удалось.
Предполагается, что "креодонты" дивергировали от эволюционной ветви ведущей к карниворам не позднее раннего палеоцена, после чего эти группы хищников эволюционировали параллельно. Современные хищные морфологически более удалены от общей предковой предковой группы, чем "креодонты". Появившись приблизительно в одно время с хищными, "креодонты" длительное время конкурировали с ними, зачастую добиваясь большего успеха благодаря быстрой дифференциации, но, в конце концов, полностью исчезли.
В Северной Америке древнейший гиенодонт — провиверрин Deltatherium — известен из первой половины среднего палеоцена (торрехонский век). Вероятно, в Азии "креодонты" появились не позднее середины палеоцена, однако древнейшие известные на сегодняшний день находки — Oxyaenidae gen. et sp. indet. из числа оксиенодонтов и Proviverrinae gen. et sp. indet. из гиенодонтов — происходят только из среднего палеоцена Монголии (свита Наран-Булак). Возможно, им синхронна древнейшие североамериканские оксиенодонты — Dipsalidictis, Dipsalodon и Tytthaena (середина тиффанского века). Таким образом, в среднем палеоцене оксиенодонты попали в Северную Америку и распространились здесь. Кроме того они, одновременно с Proviverrinae, в раннем эоцене достигли Европы (Palaeonictis и Prototomus соответственно, зона МР-7), но в Африку не проникли. В то же время, гиенодонты (Boualitomus, Lahimia) достигли севера Африки еще в среднем палеоцене (зеландский век).
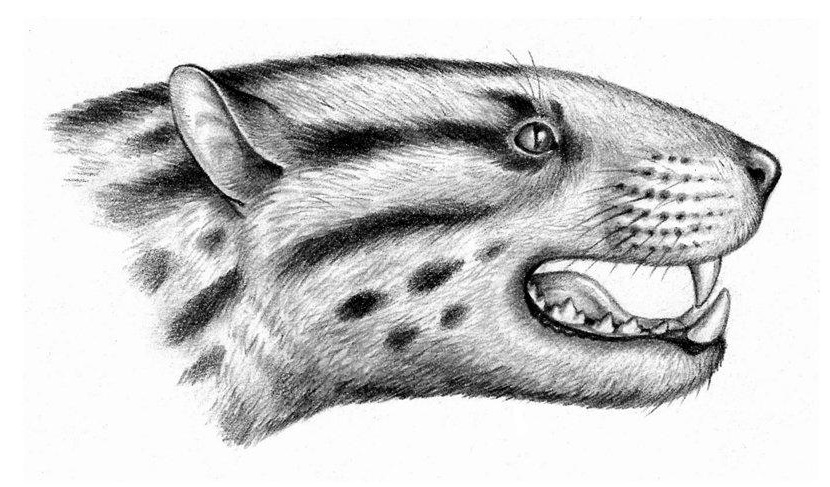
Портретная реконструкция оксиены (Oxyaena lupina). Автор — А. Кац.
Появление "креодонтов" в Европе и Северной Америке происходило синхронно с появлением млекопитающих других отрядов. Так, с оксиенодонтами в палеоцене и раннем эоцене в Северную Америку из Азии вселяются представители Mesonychia, Rodentia, Primates, Artiodactyla. Переселение в Азию из Северной Америки происходило в меньших масштабах, что, вероятно, объясняется меньшей конкурентоспособностью североамериканской фауны в раннем палеогене.
В Азии оксиенодонты представлены палеоценовым "Охуаеnа" sp. и специализированным гигаетским родом Sarkastodon (средний эоцен). Архаичное плантиградное строение автоподия оксиенодонтов, по-видимому, предопределило их меньшую конкурентноспособность по сравнению с дигитиградными гиенодонтами и мезонихиями. Можно полагать, что, сформировавшись в Азии, оксиенодонты не были здесь многочисленны в палеоцене. Только после вселения в Северную Америку группа смогла освоить широкий спектр экологических адаптаций. В раннем эоцене (зона МР 9) европейские оксиенодонты вымерли.
Первый пик разнообразия "креодонтов" отмечен в раннем эоцене (уосатчский и бриджерский века) и связан с переседение из Азии в Северную Америку оксиенодонтов и части гиенодонтов (Limnocyoninae и Proviverrinae). В начале среднего эоцена обособляются Machaerodinae, эндемичны для Северной Америки. К середине бриджерия Limnocyoninae исчезают. К концу века вымерли азиатские и американские оксиенодонты, что привело к мощному всплеску разнообразия гиенодонтов.

Портретная реконструкция апатэлюра (Apataelurus kayi). Автор — А. Кац.
Появление Pterodontinae произошло в начале среднего эоцена в Азии (Isphanatherium, Neoparapterodon sp., нижнеалайская подсвита). Pterodontinae вселились в Северную Америку из Азии в начале среднего эоцена, предположительно не позднее середины уинтского века, до момента прекращения существования берингийского моста. Из Америки подсемейство по северо-атлантическому мосту проникло в Европу, где его появление фиксируется в зоне MP 11 Гейзейталя (Oxyaenoides). В Африке Pterodontinae известны начиная с позднего эоцена (Apterodon).
В Азии Hyaenodontinae появляются во второй половине среднего эоцена (формация Шара-Мурун); в Северной Америке — в конце среднего эоцена (начало дюшенского века); в Европе Hyaenodontinae известны с зоны MP 17 (поздний эоцен), куда они вселяются из Азии. В Африке находки подсемейства не известны. В конце среднего - начале позднего эоцена происходит бурная адаптивная радиация Hyaenodontinae. Только в раннем чадроне Северной Америки известно 8 их видов. В Европе обитало 11 видов. В позднем эоцене наблюдается вспышка разнообразия Pterodontinae, из них описано 8 родов в Европе, 6 родов в Азии и 5 родов в Африке (в позднем олигоцене - миоцене отсюда в общей сложности известно 7 родов), в Северной Америке — 2 рода.
С середины эоцена среди гиенодонтов появляются животные, способные охотиться на достаточно большую добычу, не уступавшую в размерах им самим. Не исключено, что это имело непосредственную связь с повсеместным сокращением численности и разнообразия древних копытных мезонихий, до этого прочно занимавших ниши крупных наземных хищников. К концу эоцена некоторые гиенодонты уже достигают гигантских размеров (Hemipsalodon, Hyaenodon).
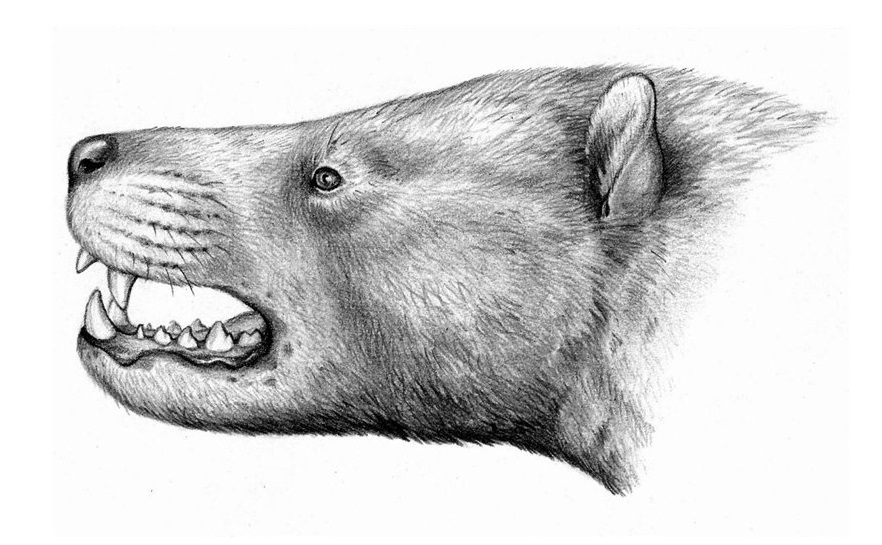
Портретная реконструкция мегистотерия (Megistotherium osteothlastes). Автор — А. Кац.
В позднем эоцене происходит угасание Proviverrinae. Они доживают до конца олигоцена - начала миоцена только в Африке и Южной Индии. Но в Африке в это время обособляются Teratodontinae, представляющие собой уклоняющееся направление эволюции "креодонтов" — специализацию в склерофагии. В раннем олигоцене вымирают все Pterodontinae Северной Америки и Евразии (помимо наиболее крупного Hyainailouros), несколько уменьшается разнообразие Hyaenodontinae.
В эоцене и на рубеже эоцена и олигоцена отмечается похолодание, в Центральной Азии нарастает аридность климата. В олигоцене изменяется спектр потенциальных пищевых объектов "креодонтов": сокращается численность и разнообразие непарнопалых, происходит радиация полорогих из числа парнопалых. В этих условиях в Северной Америке в конце олигоцена "креодонты" полностью вымирают. В Европе до начала миоцена (зона MN 2) доживает только Hyainailouros, а в Азии — крупные виды Hyaenodon (горизонт соответствует европейским зонам MN 3 и 4) и гигантский Hyainailouros. Дольше всего они удерживаются в Индии и Восточной Африке (датировки калий-аргоновым методом дают 14-11 млн. лет), откуда происходят находки последних, наиболее крупных представителей родов Megistotherium, Hyainailouros и Dissopsalis.
Образ жизни
"Креодонты" занимали экологические ниши наземных хищников всех размерных классов и играли ключевую роль в биоценозах как консументы второго порядка, некоторые из них могли быть падальщиками. Многочисленные представители отряда предвосхитили большинство экологических групп настоящих хищных (Carnivora). Среди них были звери, аналогичные крупным и мелким куньим (включая каланов), псовым, виверрам, мангустам, енотам и медведям. Однако многие "креодонты" настолько своеобразны, что не похожи ни на одного другого хищника (например, гигантский Sarcastodon с очень широким и коротким лицевым отделом).
В большинстве своем группа включала наземных животных, хотя некоторые ее ранние представители неплохо лазали по деревьям или даже вели вполне древесный образ жизни (например, куницеобразный Lesmesodon). Быстро бегать, да еще и на приличные расстояния, ранние "креодонты" (Oxyaenidae, Proviverrinae, Limnocyoninae) не могли. По сути, они передвигались не быстрее человека и только на короткие дистанции. Охотилось большинство ранних "креодонтов" преимущественно из засады, подкарауливая зазевавшуюся добычу. Однако со временем ряд "креодонтов" (Hyaenodontidae) становится активными бегающими хищниками открытых пространств. Значительную скорость эти животные развивать не могли, но, вероятно, были очень активными и выносливыми охотниками. Кроме того, из числа "креодонтов" выдвинулись и саблезубые формы (Machaeroidinae), специализированные к охоте на какую-то вполне определенную добычу. Примечательно также, что по меньшей мере один представитель отряда (склерофаг Teratodon) освоил водную среду обитания.
Вымирание
Видимо, основными причинами исчезновения "креодонтов" являлись консервативность их морфологии, ее приверженность к архаичным чертам, которые представители отряда пронесли на протяжении всей истории его существования, и их крайняя специализация как хищников, жестко привязанных к определенным условиям и добыче. Все это способствовало сужению адаптивных возможностей в постоянно меняющемся мире, а значит, ставило под вопрос само существование группы.
В частности, одной из возможных причин вымирания мог стать проглядывающийся эволюционный тупик в развитии архаичного мозга "креодонтов". У них, в отличие от эоценовых карнивор, имелся рисунок кольцевых структур из борозд и извилин, спереди от структуры, называемой "медвежьим" ромбом. Такое расположение могло блокировать дальнейшее усложнение коры мозга. Возможно, именно это завело "креодонтов" в тупик и не дало на равных соперничать с настоящими хищными.
Зачастую неверно утверждается, что "креодонты" были вытеснены более адаптивными карниворами. Однако согласно данным палеонтологической летописи, почти все группы настоящих хищных (за исключением мелких, древесных и некоторых узко специализированных форм) получают широкое распространение только некоторое время спустя после исчезновения "креодонтов". Очевидно, различные "креодонты" раньше настоящих хищных заняли большинство ниш наземных хищников и вплоть до своего вымирания просто не допускали в них потенциальных конкурентов.
Ссылки
Евсеев Р. Creodonta: история альтернативных хищников. — 2011 (Иные миры) перейти
Литература
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 3 — М.: Мир, 1993
Лавров А.В. Креодонты Азии (Creodonta, Mammalia): морфология и систематика. — М., 1999
Лавров А. В. Неизвестные хищники креодонты. ПалеоМир 1 (8), 2010
Основы палеонтологии. В 15 т. Т. 13: Млекопитающие / Гл. ред. Орлов Ю. А. — М.: Государственное научно-техническое издательство литературы по геологии и охране недр, 1962
Ромер А. Ш. Палеонтология позвоночных. — М. — Л.: Государственное научно-техническое издательство нефтяной и горно-топливной литературы, 1939
Россолимо О. Л., Павлинов И. Я., Крускоп С. В., Лисовский А. А., Спасская Н. Н., Борисенко А. В., Панютина А. А. Разнообразие млекопитающих. Ч. 1-3. — М.: Изд-во КМК, 2004
Савельев С. В., Лавров А. В. Окаменевшие мозги. Химия и жизнь 1, 2002
Evolution of Tertiary Mammals of North America. Vol. 1: Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals. Edited by Janis C. M., Scott K. M. & Jacobs L. L. — Cambridge: Cambridge University Press, 1998
Kemp T. S. The Origin and Evolution of Mammals. — Oxford: Oxford University Press, 2005
Macdonald D. W. The Velvet Claw. A Natural History of the Carnivores. — New York: Parkwest; London: BBC Books, 1993
Matthew W. D. Additional Observations on the Creodonta. Bulletin of the AMNH 14 (1), 1901
Rose K. D. The Beginning of the Age of Mammals. — Oxford: Johns Hopkins University Press, 2006