Лептиктиды — Leptictida
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Отряд †Лептиктиды (Leptictida)
Лептиктиды (Leptictida MсKenna, 1975) — отряд клады эвтериев (Eutheria Gill, 1872), происходящий из позднего мела - раннего олигоцена Северной Америки, раннего - позднего эоцена Европы и позднего палеоцена - раннего эоцена Азии. Установлены 3 семейства с приблизительно 17 родами.

Разнообразие и относительные размеры лептиктид (Leptictida). Автор — artbyjrc.
Родственные связи
Установление родственных связей лептиктид с другими группами млекопитающих сильно затруднено. Разые исследователи сближали их с ежообразными (Erinaceomorpha), тупайями (Scandentia), плезиадапиобразными (Plesiadapiformes), приматами (Primates), грызунами (Rodentia), прыгунчиками (Macroscelidea), апатемиевыми (Apatemyidae), пантолестовыми (Pantolestidae), палеанодонтами (Рalaeanodontа) и некоторыми другими. Подобный разброс мнений обусловлен прежде всего архаичностью строения черепа и зубов лептиктид, базового для эвтериев. Примитивностью морфологии объясняется и то, что на протяжении 1960-х и 1970-х гг. они зачастую считались связующим звеном между различными группами ранних плацентарных, включая и приматов. Сегодня принято считать, что многие общие с другими примитивными эвтериями признаки лептиктид объясняются плезиоморфией и на самом деле это достаточно самостоятельная в эволюционном смысле группа.
В настоящее время лептиктиды обычно рассматриваются в качестве одной из древнейших ветвей, отделившихся от базальных эвтериев (Еutheria) еще в меловое время. Иногда их происхождение связывают с залямбдалестидами (Zalambdalestida) и желестидами (Zhelestida). Все они являются представителями позднемеловой радиации стволовых неплацентарных эвтериев, предшествовавшей и сосуществовавшей с кроновой группой ранних плацентарных (Placentalia). Представители обеих волн эволюции эвтериев выиграли от вымирания нептичьих динозавров в конце мела, но в течение палеогена постепенно сдали позиции более продвинутым групам плацентарных.
Возможно, из ныне живущих групп к лептиктидам в родственном отношении наиболее близки насекомоядные (Eulipotyphla). Об этом свидетельствуют многочисленные общие черты краниодентальной анатомии, хотя между двумя этими таксонами имеются и существенные морфологические различия (например, в строении глазницы и слуховой области). Согласно кладистическим исследованиям, лептиктиды могут иметь общего предка с эвархонтоглиресами (Euarchontoglires). Значительная длина нижних отделов конечностей в сравнении с верхними отделами, что особенно хорошо заметно на задней их паре, указывает на происхождение от быстробегающих животных.
Систематика
Весьма неопределенно и таксономическое положение самих лептиктид. Чаще всего они рассматриваются в качестве отряда, реже — надотряда или подотряда. Большинством авторов на сегодняшний день собственно лептиктиды принимаются в качестве отряда с как минимум одним валидным семейством — лептиктовыми (Leptictidae), объединяющими типичных палеогеновых представителей группы. Помимо них, в отряд могут дополнительно включаться семейства гипсониктоповых (Gypsonictорidae), кульбекиевых (Kulbeckiidae), дидимоконовых (Didymoconidae), псевдоринкоционовых (Pseudorhyncocyonidae) (последние нередко рассматриваются в качестве подсемейства в составе Leptictidae), а также отдельные роды, не отнесенные к тому либо другому семейству. Таким образом, не исключается парафилетичность лептиктид в их нынешнем понимании.
Здесь принимается следующий состав отряда:
семейство †Гипсониктоповые (Gypsonictopidae Van Valen, 1967);
семейство †Лептиктидовые (Leptictidae Gill, 1872);
семейство †Псевдоринкоционовые (Pseudorhyncocyonidae Sigé, 1974).
Возраст и распространение
Древнейшие представители группы известны из позднего мела (кампанский ярус) Северной Америки. Возникнув на этом континенте, лептиктиды сохраняют здесь сравнительное разнообразие на протяжении всего палеоцена и эоцена и исчезают только в раннем олигоцене (уайтский ярус). Несколько родов также описаны из временного промежутка с раннего по поздний эоцен Европы (ипрский - приабонский ярусы) и позднего палеоцена - раннего эоцена Азии.
Строение и функции
Морфоанатомия некоторых лептиктид довольно хорошо изучена, что позволяет с достаточной уверенностью судить о всей группе. Ее представители демонстрируют архаичный план строения с многими характерными для ранних эвтериев признаками, которые сочетаются с рядом специализированных адаптаций.
Размеры небольшие: общая длина тела составляет 40-90 см, а вес оценивается в 0,2-2 кг.

Скелет лептиктидия (Leptictidium auderiense) в блоке породы из Мессельского карьера в земле Гессен, Германия. Национальный музей естествознания, Париж.
Черепные пропорции лептиктид в общих чертах сходны с таковыми у тупай и прыгунчиков, но в деталях краниальные характеристики этих групп существенно различаются. Лицевой отдел черепа длинный. Носовые кости удлинены и образуют конический рострум, который несет ямки, борозды и гребешки, служащие для крепления мышц рыльца. Вероятно, у многих лептиктид имелся подвижный хоботок. Эти животные обладали хорошо развитым обонянием, их носовые ходы расширены и занимают в черепных костях немало места. Заглазничный отросток отсутствует. Иногда имеется сагиттальный гребень. Мозговая камера довольно длинная. Барабанная полость отделяется от твердого неба значительным промежутком, как у тупай. Лептиктиды уникальны среди эвтериев наличием заметного треугольного выступа теменной кости на ее затылочной поверхности.
Примитивность черепа проявляется в удлиненной морде и сохранении цельной, длинной и тонкой скуловой дуги (в отличие от многих настоящих насекомоядных). Сонная артерия проходит медиально. К продвинутым признакам относится включение в слуховой барабан внутренней барабанной кости, которая, впрочем, не образует ее сплошного покрытия. Таким образом, слуховой барабан неполный и открытый снизу. Барабанная перепонка поддерживалась кольцевидной барабанной костью, как у большинства современных землероек. Костного слухового канала нет. Мозг весьма примитивен: обонятельные луковицы очень крупные при сравнительно коротком мозжечке.
Зубная формула: I3-2/i3-2, C1/c1, P5-4/p5-4, M3/m3. I1 и i1 у многих родов отсутствуют. I1 и i2 увеличены и направлены вперед. Клыки разной величины. Клыки и Р1/р1 однокоренные. Предкоренные простой конусовидной формы. У многих лептиктид наблюдается утрата P3 (центрального предкоренного). Последние предкоренные почти всегда моляризованы, верхние — с двумя наружными и двумя внутренними бугорками. Коренные примитивного режущего типа с высокими, острыми бугорками. Верхние коренные в ширину больше, чем в длину, треугольные, с V-образным рисунком, образованным пара- и метаконом, расположенными близко к наружному краю коронки, и протоконом, соединенным гребешками с пара- и метастилем; часто с зачаточным гипоконом и иногда з зачаточными прото- и метаконулем. Нижние коренные с высоким трехзубчатым тригонидом, сдавленном в передне-заднем направлении, и низким талонидом, на котором находится обширный мелкий бассейн.
Посткраниальный скелет лептиктид также изучен достаточно хорошо, известны полные скелеты представителей нескольких родов. Шея и туловище умеренной длины. Передние конечности укорочены, но при этом довольно сильные. Плечевая кость с внутренним надмыщелковым отверстием. Лучевая и локтевая кости относительно тонкие, локтевой отросток короткий. Задние конечности значительно удлинены, это особенно характерно для тонкого бедра с небольшим III-м вертелом, большой берцовой кости и стопы; предплюсна и метатарзальный отдел удлинены умеренно. У ранних лептиктид большая и малая берцовые кости иногда разделены, у типичных представителей группы сращены. Хвост длинный, может насчитывать более 40 позвонков.
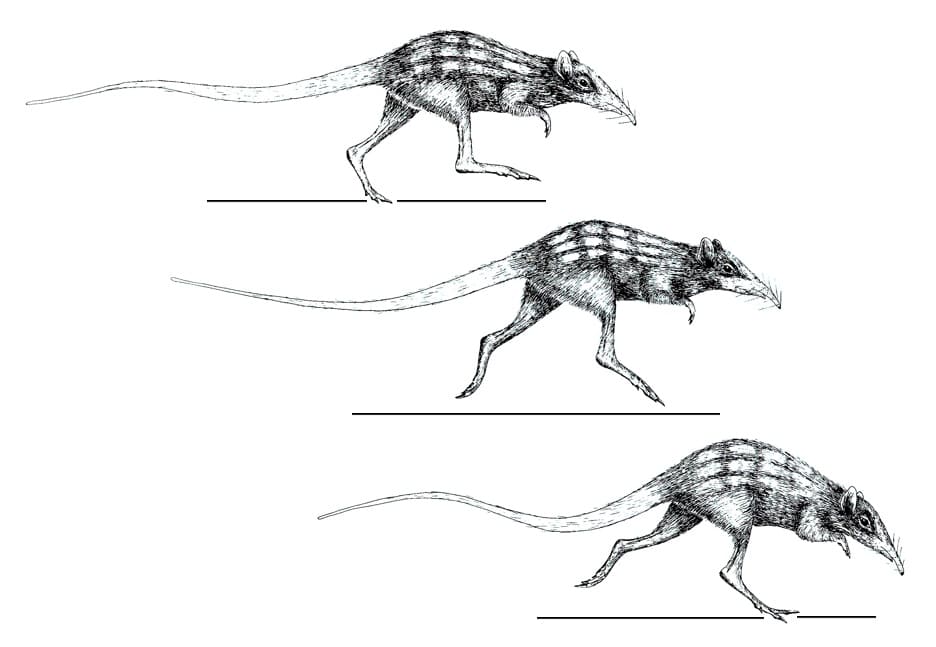
Строение посткраниального скелета свидетельствуют о том, что лептиктиды являлись наземными животными, передвигающимися на всех четырех или только на двух задних конечностях. Двуногое передвижение могло осуществляться посредством бега или рикошетирующих прыжков. Скорее всего, это были именно прыжки, поскольку бег на двух ногах требует гораздо больших энергетических затрат и попросту был бы невыгоден таким мелким животным. Хвост при этом служил для поддержания равновесия. Морфология передних конечностей указывает на то, что лептиктиды рыли ими землю, добывая пищу или устраивая убежища. Устройство зубочелюстного аппарата свидетельствует о их насекомоядности и всеядности. Из современных млекопитающих лептиктиды экологически более всего напоминали прыгунчиков (Macroscelidea) и бандикутообразных (Peramelemorphia).


Гипотезы локомоции лептиктид на примере лептиктидия (Leptictidium nasutum): вверху — двуногий бег, внизу — рикошетирующие прыжки.
Вымирание
Лептиктиды появились и процветали в теплых и влажных условиях лесных экосистем. Исчезновение этих животных связывают с прогрессирующим похолоданием и иссушением климата, что вызвало разрежение лесных массивов и сокращение занимаемых ими площадей. В итоге лептиктиды не смогли успешно противостоять новым конкурентам и хищникам открытых пространств. Их численность постепенно уменьшалась, пока не последовало неизбежное вымирание. Потомков после себя лептиктиды не оставили.
Ссылки
Leptictida (Paleobiology Database)
перейти
Литература
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 3 — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 13: Млекопитающие / Гл. ред. Орлов Ю. А. — М.: Государственное научно-техническое издательство литературы по геологии и охране недр, 1962
Agusti J., Anton M. Mammoths, sabertooths, and hominids: 65 million years of mammalian evolution in Europe. — New York: Columbia University Press, 2002
Evolution of Tertiary Mammals of North America. Vol. 2: Terrestrial Carnivores, Ungulates, and Ungulatelike Mammals. Edited by Janis С. М., Gunnell G. F. & Uhen M. D. — Cambridge University Press: Cambridge, United Kingdom, 2008
Kemp T. S. The Origin and Evolution of Mammals. — Oxford University Press: Oxford, 2005
Rose K. D. The Beginning of the Age of Mammals. — Johns Hopkins University Press: Oxford, 2006