

Находка окаменелостей лисовиции была совершена на территории кирпичного завода в селе Лисовице (ныне данная его часть выделена в деревню Липе-Сленске), которое находится к западу от города Люблинец на юго-западе Польши. Первыми на фрагменты костей наткнулись рабочие в яме, откуда они брали глину для производства кирпича. Об этом стало известно геологу-любителю Роберту Борзецки, который в 2005 г. поставил в известность профессиональных ученых. Первичная разведка подтвердила существование окаменелостей в глинистой толще. Организованные палеонтологические работы были начаты в 2007 г. и продолжались до 2017 г. За это время было обнаружено более тысячи костей и их фрагментов, принадлежащих некогда обитавшему здесь животному сообществу.




Первое сообщение в научной прессе об открытии гигантского зверообразного сделали в 2008 г.палеонтологи Ежи Дзик, Томаш Сулей и Гжегож Недзведзкий. Из-за крупных размеров специалисты сначала приписали окаменелости завроподу, но когда нашлись фрагменты черепа и кости конечностей выяснилась их принадлежность дицинодонту. Таксон Lisowicia bojani был установлен в 2019 г. Сулеем и Недзведзким. Его родовое наименование происходит от села Лисовице, а видовой эпитет присвоен в честь Людвига Генриха Боянуса, жившего в XVIII в. естествоиспытателя.
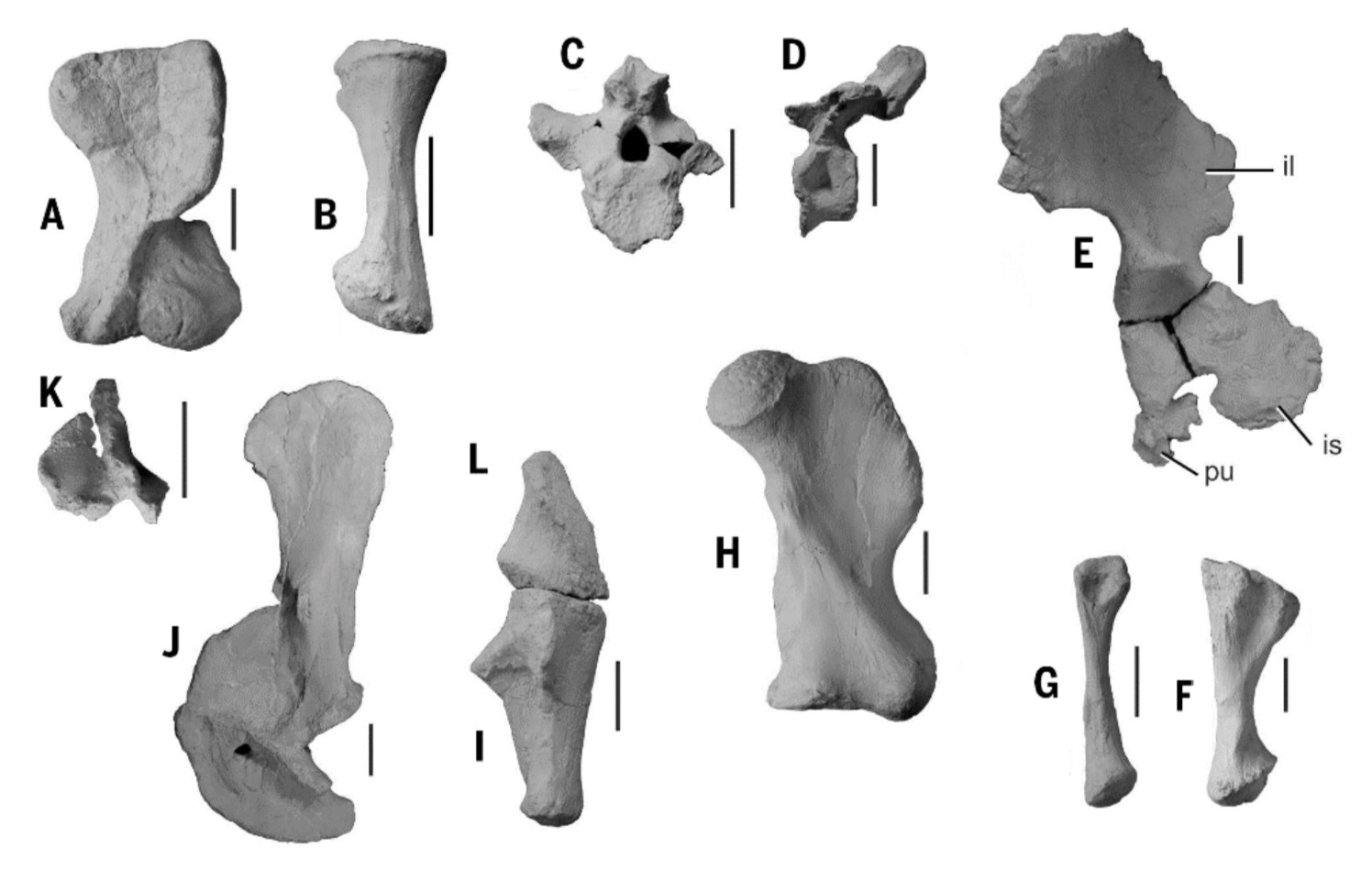
Найденные остатки лисовиции представляют собой фрагменты черепа и большинства элементов посткраниального скелета, происходящих от нескольких особей сходной величины. В качестве голотипа выбрана левая плечевая кость (образец ZPAL V.33/96). Таким образом, лисовиция является первым позднетриасовым европейским дицинодонтом, известным по большему количеству ископаемого материала, чем отдельные изолированные кости.
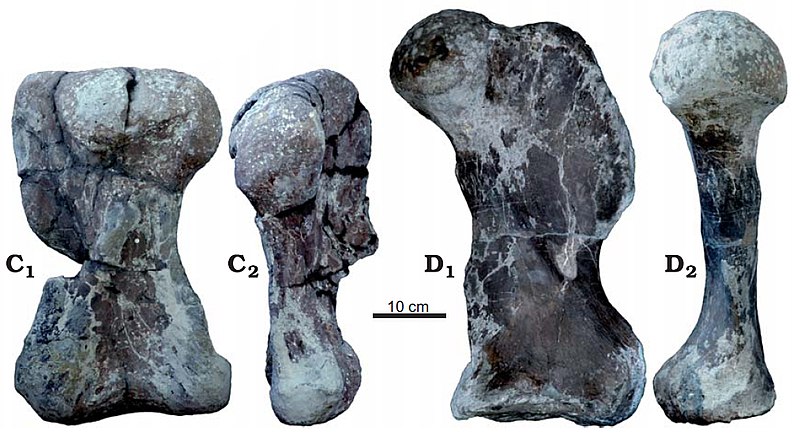
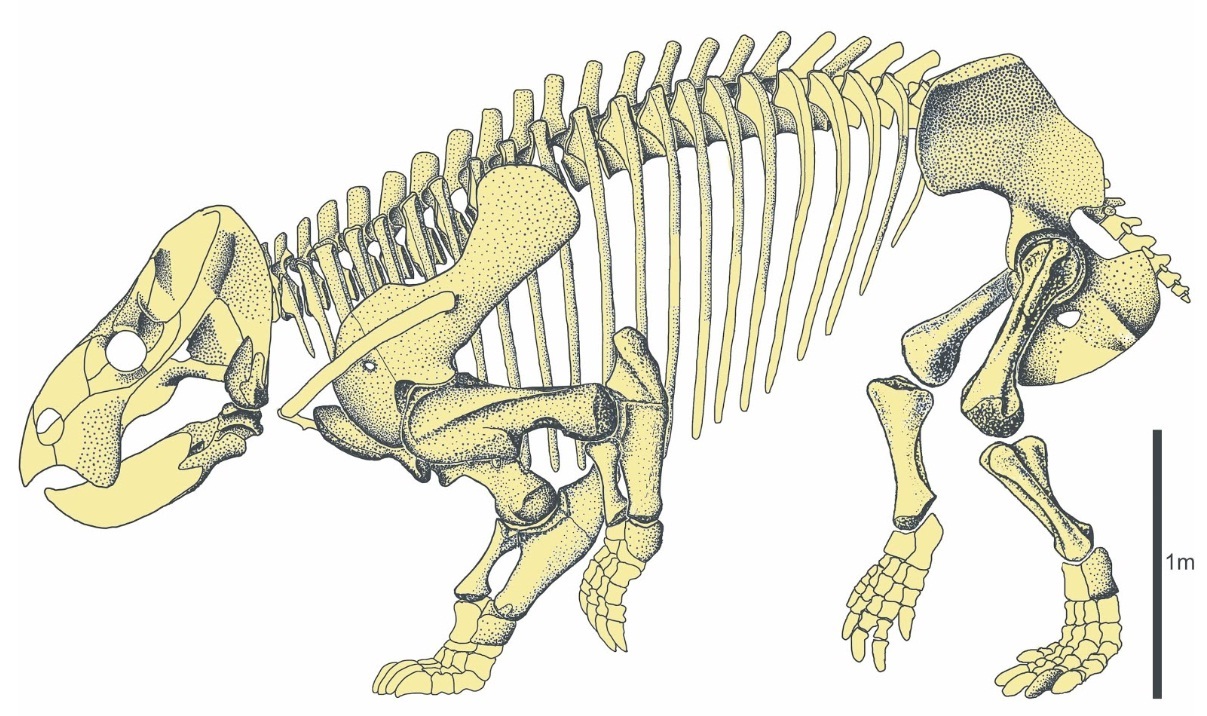


До описания лисовиции не были известны триасовые дицинодонты, расчетная длина тела которых превышала 3-3,5 м, а масса 1-2 т. Таким образом, крупнейшие из них по своим габаритам не превосходили самых больших пермских диноцефалов. В конце триаса больших размеров, значительно превышающих таковые дицинодонтов, начали достигать динозавры из группы завроподообразных, включая завропод и их ближайших родственников; в начале юрского периода некоторые из них уже достигали свыше 15 м в длину и весили более 10 т. Открытие лисовиции предполагает, что возможность достижения динозаврами значительной величины не была обусловлена только их анатомией, а большие размеры могут являться результатом взаимодействия ряда экологических факторов, также вызвавших увеличение общих размеров представителей других групп животных — например, дицинодонтов.

Открытие лисовиции также показывает, что, вопреки более ранним представлениям, эволюционная радиация ранних завроподообразных не совпала с исчезновением дицинодонтов из палеонтологической летописи, а крупные дицинодонты сосуществовали в позднем триасе с большими завроподообразными.

Судя по всему, лисовиция держалась по берегам водоемов и вела земноводный образ жизни. Судя по сопутствующим окаменелостям, в ее рацион могли входить хвойные из родов Brachyphyllum, Pagiophyllum и Hirmeriella. В найденных поблизости копролитах, приписываемых этому дицинодонту, выявлены пыльца, кутикулы и частицы древесины голосеменных растений. Это указывает на высокую питательную ценность потребляемой растительности. На лисовицию, вероятно, охотился крупный (расчетная длина 5-6 м) архозавр смок (Smok wawelski), известный из того же местонахождения. Некоторые обнаруженные кости лисовиции несут следы укусов, соответствующие его зубам.

Ссылки