Лофиалеты — Lophialetes
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Клада Скротиферы (Scrotifera)
Клада Феревунгуляты (Fereuungulata)
Клада Эвунгуляты (Euungulata)
Клада Мезаксонии (Mesaxonia)
Отряд Непарнопалые (Perissodactyla)
Подотряд Тапирообразные (Tapiromorpha)
Инфраотряд Носорогообразные (Ceratomorpha)
Надемейство Тапироиды (Tapiroidea)
Семейство †Лофиалетовые (Lophialetidae)
Род †Лофиалеты (Lophialetes)
Вид †Lophialetes expeditus
Лофиалеты (Lophialetes Matthew et Granger, 1925) — монотипический род семейства лофиалетовых (Lophialetidae Matthew et Granger, 1925), происходящий из среднего эоцена Азии.
Реконструкция внешнего облика лофиалета (Lophialetes expeditus). Автор — Р. Учитель.
Синонимы: Lophiodon Marsh, 1871 (частью); Hyrachyus Leidy, 1871 (частью).
Единственный установленный вид — Lophialetes expeditus Matthew et Granger, 1925 (тип). Его остатки обнаружены в автономном районе Внутренняя Монголия и провинции Юньнань, Китай, а также Овор-Хангайском аймаке, Монголия. Они датируются средним эоценом (аршантский - ирдынмангский ярусы).
Остатки, определенные только до рода и обозначенные Lophialetes spp., происходят из среднего - позднего эоцена Казахстана, Киргизии, Монголии и Китая (Синьцзян-Уйгурский автономный район, автономный район Внутренняя Монголия, провинции Хэнань и Шаньдун).
История изучения
Род и вид Lophialetes expeditus установили в 1925 г. Уильям Диллер Мэтью и Уолтер Уиллис Грейнджер по верхнечелюстному фрагменту из Ирдын-Манга во Внутренней Монголии. Некоторое описание черепа и ряда посткраниальных элементов (главным образом кисти и стопы) дал в 1965 г. Л.В. Радинский. Череп и посткраниальный материал в 1979 г. детально описал В.Ю. Решетов.
Ископаемые остатки Lophialetes expeditus обильны в местонахождениях Ирдын-Манга, Сичуань и Лулян (Китай), Улан-Шире, Шара-Мурун, Хайчин-Ула II, Хайчин-Ула III, Холболджи-Нур I и Кэмп-Маргетс (Монголия), Тору-Айгыр (Киргизстан). В сборах совместной советско-монгольской экспедиции имеются многочисленные остатки этого тапироида: разрозненные зубы, челюстные фрагменты, черепа, части посткраниального скелета.
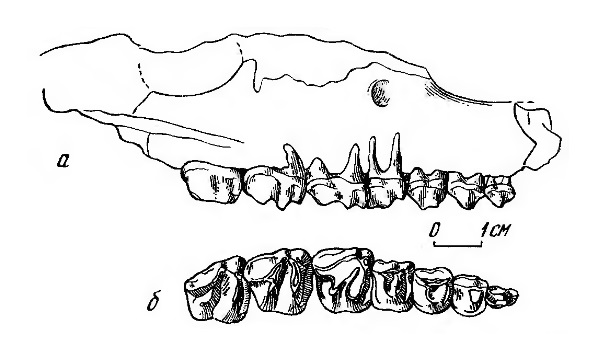
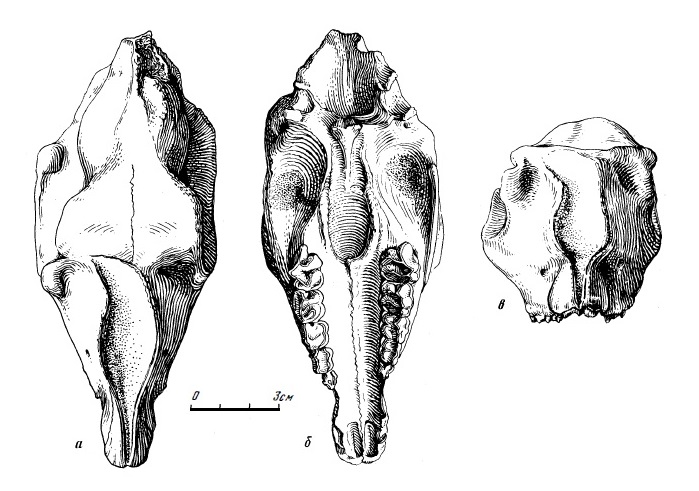
Типовой образец лофиалета (Lophialetes expeditus) — верхнечелюстной фрагмент с Р4-М3 из Ирэн-Дебасу, Китай: снаружи (а), жевательная поверхность верхних щечных зубов (б).
На сегодняшний день существование других видов рода, формально описанных как Lophialetes minutus и Lophialetes tokunagai сомнительно. Lophialetes tokunagai, выделенный в 1939 г. Ф. Такаи по нижней челюсти с зубами из Хосана в Корее, по мнению Л.В. Радинского, не тапироид, а какой-то род носорогообразных. Вероятно, другому животному принадлежат и остатки Lophialetes minutus из позднего эоцена провинции Хэнань, Китай. Неопределенным является положение Lophialetes yunanensis, описанного в 1982 г. из Луньана, Китай; вероятно, это младший синоним Lophialetes expeditus.
Ископаемые остатки лофиалета (Lophialetes expeditus) из отложений Хайчинской свиты местонахождения Хайчин-Ула III, Монголия.
Строение и функции
Размеры Lophialetes средние для группы: длина его тела составляла 80-100 см, высота в холке — 50-60 см, а вес оценивается в 20-40 кг. Это было легко сложенное животное, не похожее на других тапироидов и отдаленно напоминающее современных антилоп, с хоботкообразной верхней губой и сильно удлиненными стройными трехпалыми конечностями.
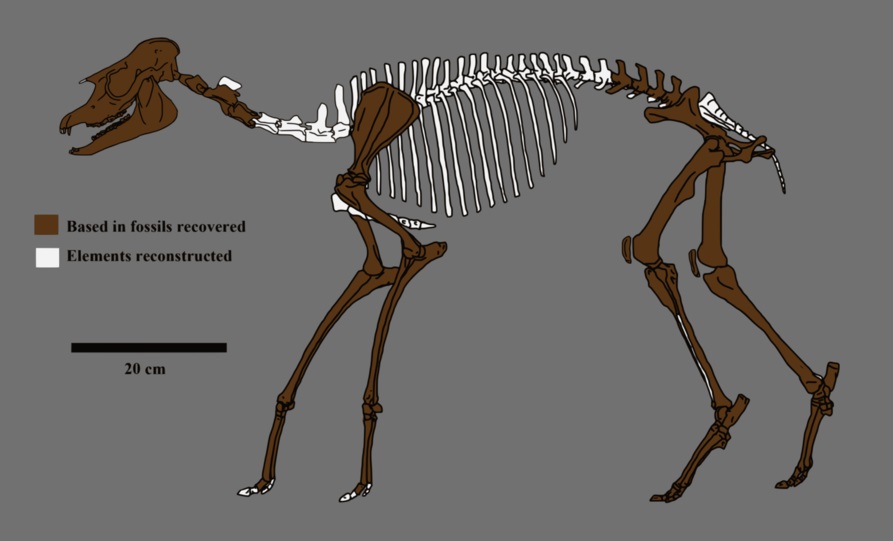
Реконструированный скелет лофиалета (Lophialetes expeditus).
Череп
Череп удлиненный, сравнительно низкий и узкий. Его длина составляет 18,4-19 см, из которых на лицевой отдел приходится в среднем 9 см. Ширина черепа по передним краям глазниц составляет 6,5 см, над диастемой — 2,2 см, над I3 — 2,3 см. Ростральная часть черепа в области диастемы сужается, резцовая область несколько расширена. Предчелюстные кости вытянутые, низкие и несросшиеся. В передней части каждая предчелюстная кость несет по 3 резца, распространяется назад до уровня Р2 , постепенно сужаясь. Максимальная высота (над Р2) — 1,4 см. Предчелюстная кость молодых особей имеет более округлые очертания.
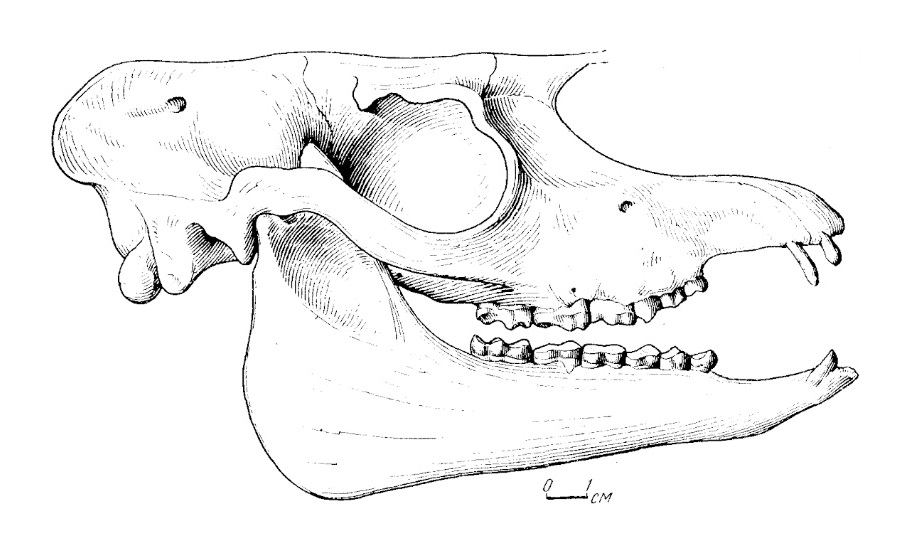
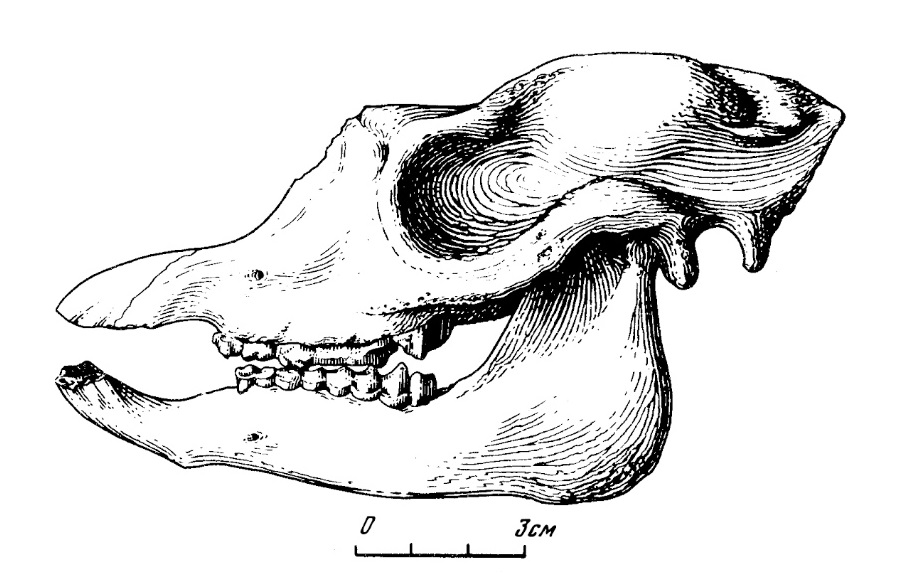
Прорисовка черепа с нижней челюстью лофиалета (Lophialetes expeditus) из отложений хайчинской свиты местонахождения Хайчин-Ула III, Монголия.
Верхнечелюстные кости относительно высокие и изогнуты вверх, что придает черепу несколько бульдогообразные очертания. Надальвеолярная часть верхнечелюстной кости сильно шероховатая, здесь ясно выражены места прикрепления жевательных и губных мышц. Височный гребень полого выступает в средней, наиболее высокой части верхнечелюстной кости, постепенно сходит на нет на уровне М1-2. Высота верхнечелюстной кости над М3 — 5,8 см. Предглазничная ямка глубокая, слегка вытянутая в верхне-нижнем направлении, расположена выше переднего края глазницы. Подглазничное отверстие расположено над передним краем M1, относительно крупное. Имеется намечающаяся продолговатая впадина — дивертикулюм, где начинается мышца подниматель верхней губы.
Прорисовка черепа лофиалета (Lophialetes expeditus) из отложений хайчинской свиты местонахождения Хайчин-Ула III, Монголия.
На образце наилучшей сохранности передние концы носовых костей обломаны. Однако, видно, что носовые кости сильно укорочены, их передние края, по-видимому, не простирались дальше переднего края Р2. Носовая вырезка широкая и далеко уходит назад, закругляясь у взрослых особей над М2. Носовые кости ориентированы несколько дорзально, что особенно подчеркивается предлобной впадиной. По линии шва, разделяющего обе носовые кости, проходит глубокая долинка. Горизонтальная и вертикальная плоскости носовой кости расположены относительно друг друга почти под прямым углом. Носовая полость сравнительно высокая и широкая, однако, ее внутренняя структура не сохранилась.
Прорисовка черепа лофиалета (Lophialetes expeditus) из отложений хайчинской свиты местонахождения Хайчин-Ула III, Монголия.
Глазницы большие, округлой формы. Слезная кость расположена на границе лицевого и мозгового отделов черепа, занимает довольно обширную область на переднем крае глазницы. Слезный отросток относительно крупный трапециевидной формы, прерывает глазницу с передне-верхней стороны. Сильно развит надглазничный отросток лобной кости, закрывающий глазницу сверху. Заглазничное сужение сильно выражено, ширина черепа в этом месте составляет всего 3 см. Скуловые дуги неширокие (максимальная ширина между крайними точками черепа — 7,9 см). Нижний край скуловой кости образует острый, шероховатый гребень. Скуловой отросток височной кости узкий и относительно короткий; он поднимается круто вверх, образуя с височной областью небольшую, но глубокую ложбину.
Прорисовка черепа с нижней челюстью молодой особи лофиалета (Lophialetes expeditus) из отложений хайчинской свиты местонахождения Хайчин-Ула III, Монголия.
Граница лицевого и мозгового отделов черепа проходит по переднему краю глазницы, который у взрослых особей находится над М3. Мозговой отдел превосходит по длине лицевой. В молодом возрасте лицевой отдел еще более укорочен по сравнению с мозговым. Черепная коробка сравнительно низкая, удлиненной формы. Мозговая полость умеренно развита, занимает объем примерно в 1,5 раза меньше, чем у собак такого же размера. Чешуи височных костей вертикально подняты, но не заходят высоко ввиду сильного развития теменной области. Височный гребень сильно развит и отграничивает снизу область распространения височной мышцы. В передней части он переходит в скуловой отросток чешуйчатой кости, в задней части сливается с затылочным гребнем.
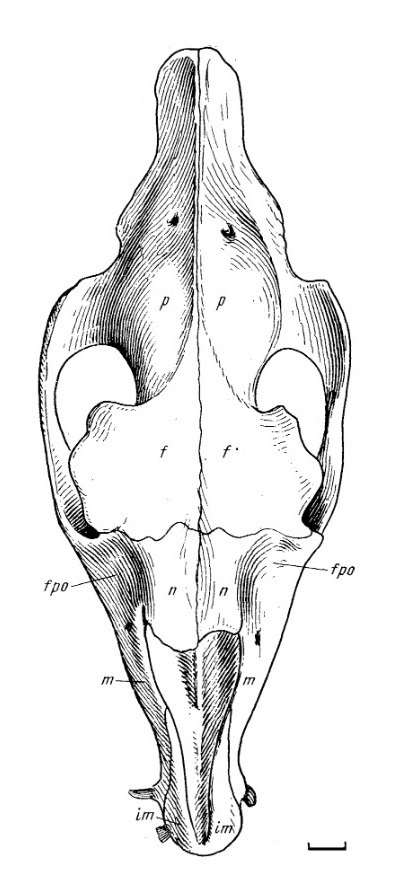
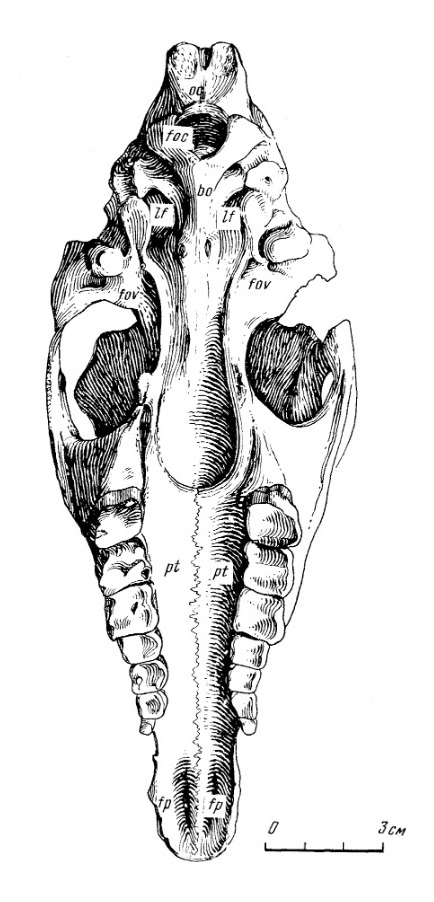
Прорисовки черепа молодой особи лофиалета (Lophialetes expeditus) из отложений хайчинской свиты местонахождения Хайчин-Ула III, Монголия: сверху (а), снизу (б) и спереди (в).
Теменные кости занимают значительное пространство, ограниченное спереди хорошо видимым лобно-теменным швом, проходящим сбоку горизонтальным височно-теменным швом и позади теменно-затылочным швом. Место соединения теменных костей образует сагиттальный гребень, увеличивающийся кзади. На верхне-задней поверхности теменной кости открывается круглое парное отверстие для сосудов и четко выраженные борозды и шероховатости для прикрепления височной мышцы. Затылочная поверхность черепа в плане напоминает треугольник с острым дорзальным углом. Затылочная кость характеризуется сильно развитым затылочным гребнем, соединяющимся сверху с сагиттальным гребнем, а сбоку с височным. Сагиттальный и затылочный гребни в профиль наклонены сильно вниз. 3атылочныe мыщелки узкие, удлиненной формы, направлены назад и вниз. Затылочное отверстие крупное. Слуховые кости полукруглые, уплощенные. Слуховое отверстие очень крупное Поверхность костного неба гладкая и плоская, почти не образующая свода. Хоаны открываются на уровне М3. Нижнечелюстная ямка относительно глубокая и узкая в передне-заднем направлении, слегка поднята вверх, в своей наружной части широкая и крупная.
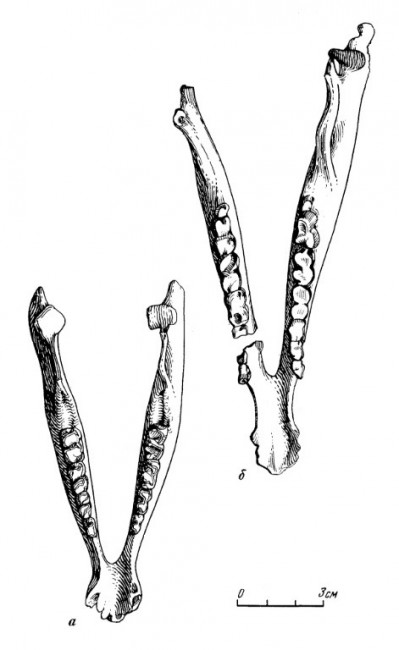
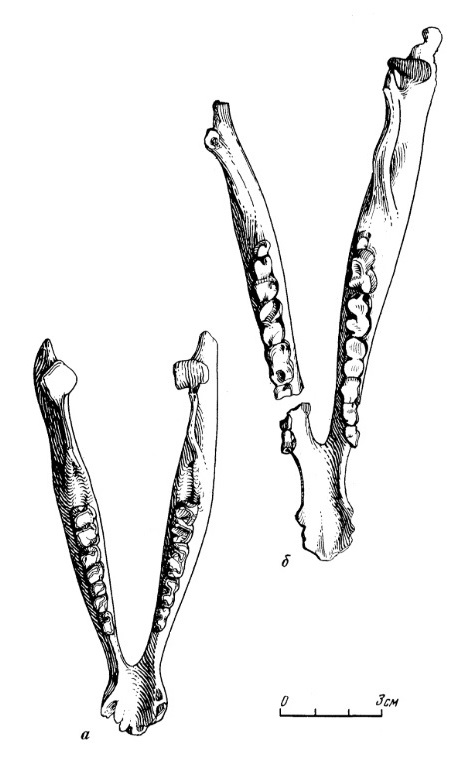
Прорисовки нижних челюстей молодой (а) и взрослой (б) особей лофиалета (Lophialetes expeditus) из отложений хайчинской свиты местонахождения Хайчин-Ула III, Монголия.
Горизонтальная ветвь нижней челюсти сравнительно низкая, постепенно снижающаяся от m3 к р1. Симфиз несколько удлинен. Диастема сравнительно длинная и узкая с расширенной симфизной частью. На уровне диастемы и под р2 расположены 2 маленьких отверстия. Восходящая ветвь широкая в основании. Задний угол челюсти округлой формы, плавно выдающийся назад по отношению к верхней части восходящей ветви. Сочленовный отросток низкий. Суставной валик удлиненный, наклонен к внутренней поверхности челюсти. Венечный отросток значительно выше сочленовного, равномерной ширины на всем протяжении и в верхней трети отклонен назад. Нижняя челюсть молодых особей имеет относительно более низкую и вздутую горизонтальную ветвь.
Зубы
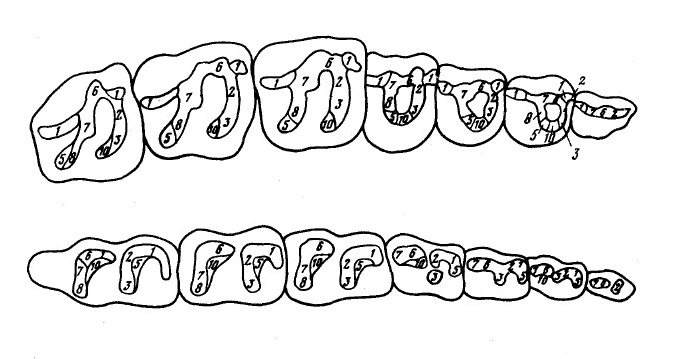
Зубная формула: І3/i3, С1/c1, Р3-4/p4, М3/m3. Верхние резцы с расширенными, уплощенными коронками. І3 не увеличен по сравнению с другими резцами. Верхний клык отделен от резцов небольшой диастемой в 4-5 мм шириной; он треугольной формы, уплощен и с острой вершинкой. Р1 треугольной формы, Р2 — субквадратной. Р3-4 немоляризованы: с одним внутренним бугром, соединенным гребнями с пара- и метаконом. M1 и М2 квадратной формы. Протолоф и металоф образуют V-образную фигуру. Эктолоф удлинен. Парастиль ниже паракона, отделен от него глубокой складкой. Метакон слегка смещен лингвально, что придает очертаниям коронки почти квадратную форму. Цингулюм хорошо развит на передней стенке М1 и М2 , а также четко выделяется в области метакона на эктолофе. Протолоф и металоф несколько скошены по отношению к эктолофу. М3 слабо сужен кзади, имея более короткую заднюю стенку сравнительно с М1-2. Металоф относительно и абсолютно короче, чем у предыдущих коренных. Заметалофная частью эктолофа длинная. Парастиль некрупный, достаточно четко обособлен. Метакон хорошо развит и уплощен.
і1-3 с расширенными уплощенными коронкам. і3 немного крупнее, чем і1 и і2. Предклыковая диастема не развита. Нижний клык треугольный, округлый в поперечном сечении. р1 у некоторых экземпляров отсутствует. Коронки р2-4 прямоугольной формы. m1-3 обладают относительно высокими и острыми протолофидами и гиполофидами. Паралофиды и металофиды удлиненной формы. На m3 развит гипоконулид. Поперечные гребни нижних коренных слабо скошены к оси зуба.
Схемы верхнего и нижнего щечных зубных рядов лофиалета (Lophialetes expeditus).
Сравнение
Череп Lophialetes еще довольно примитивен и во многом напоминает таковой более древних непарнопалых, например тапироида Нерtоdоn из раннего эоцена Северной Америки. При этом череп демонстрирует черты специализации, отличающие его от первоначального примитивного типа, характерного для всех ранних непарнопалых, таких как Hyracotherium и Heptodon. В первую очередь, эти отличия относятся к лицевому отделу — у Lophialetes он укорочен (у Heptodon лицевой отдел превосходит по длине мозговой), носовые кости сильно редуцированы по сравнению с таковыми у Heptodon и Hyracotherium (у Heptodon носовые кости простираются вперед до уровня симфиза и контактируют с предчелюстными). Соответственно увеличена и носовая вырезка: в то время, как у Heptodon задний край носовой вырезки расположен над Р1, то у Lophialetes — над М2. У Lophialetes сильнее развита предглазничная ямка. Также различно у сравниваемых форм строение затылочного отдела. Сагиттальный и затылочный гребни у Lophialetes развиты в большей степени, чем у Heptodon. Оба гребня у первого сильнее отклонены вниз. Расположение черепных отверстий у Lophialetes близко к таковому у Heptodon. Однако оно отличается от их расположения у наиболее древних представителей отряда непарнопалых. Так, у Hyracotherium в задней части глазницы находится единое крупное отверстие для прохождения всех глазничных нервов.
Lophialetes отличаются от Schlosseria более специализированными коренными зубами, более укороченными носовыми костями и соответственно увеличенной носовой вырезкой. Отличие от Eoletes состоит в строении зубной системы, в особенности М3, который у Eoletes субтреугольной формы с сильно редуцированной заметалофной частью эктолофа. Более короткие носовые кости и увеличенная носовая вырезка также отличают Lophialetes от Eoletes. От Kalakotia и Aulaxolophus рассматриваемый род отличается наличием язычного воротничка на нижних резцах, большей степенью моляризации молочных предкоренных, более скошенными по отношению к эктолофу поперечными гребнями, менее разделенными парастилем и параконом на коренных зубах, относительно менее развитым гипоконулидом на m3.
Мышцы головы
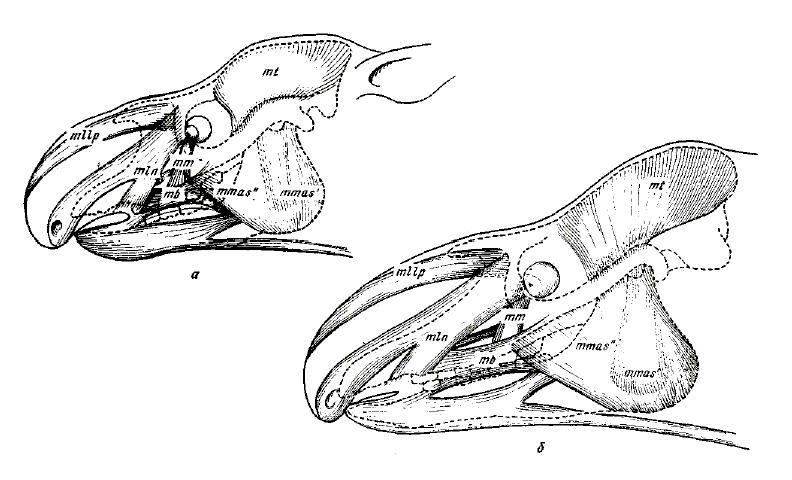
Относительно хорошая сохранность черепов Lophialetes позволяет достаточно достоверно проследить на нем места крепления лицевых мышц. Обращает на себя внимание наличие хорошо выраженной предглазничной ямки. В различных группах копытных она служит местом отхождения носогубной мускулатуры, а также является вместилищем дивертикулюма — хрящевого мешочка, открывающегося в носовой канал, а также предглазничной железы у парнокопытных. У ныне живущих Tapirus предглазничная ямка имеет вытянутую, желобообразную форму и несет двоякую функцию. Она вмещает дивертикулюм, а по ее внешнему краю крепится фасция поднимателя верхней губы. Предглазничная ямка Lophialetes несколько вытянутой формы, но не столь глубокая, как у Tapirus, и, судя по обилию отпечатков мышц на ее стенках, здесь отходил подниматель верхней губы. В то же время вполне вероятно, что дивертикулюм у Lophialetes мог быть уже значительно развит (судя по объему носовой полости и увеличенной носовой вырезке) и частично заходил в предглазничную ямку.
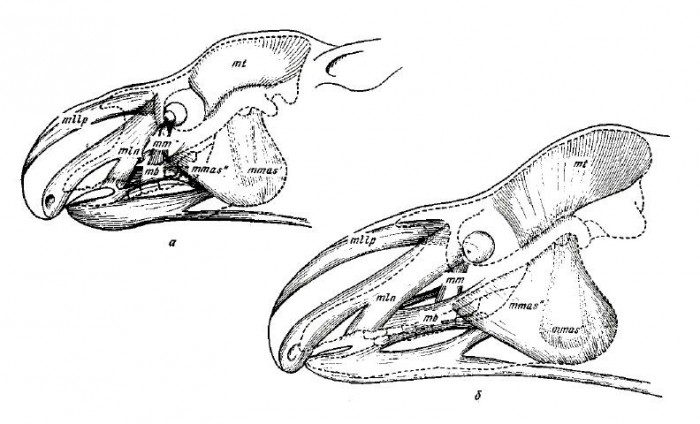
Реконструкция мышц головы молодой (а) и взрослой (б) особей лофиалета (Lophialetes expeditus).
Шероховатость на переднем крае глазницы говорит о наличии сильной носогубной мышцы. Бугристая поверхность над альвеолярной областью указывает на прикрепление губной мышцы. Все эти данные свидетельствуют о значительной подвижности мускулатуры лицевого отдела черепа Lophialetes, в особенности мышц, связанных с движениями верхней губы и носа. Это, в свою очередь, позволяет предположить подвижность хоботкообразной верхней губы. Следует отметить, что среди современных непарнопалых помимо тапиров чрезвычайно подвижной верхней губой обладает современный черный носорог (Diceros bicornis). Однако у носорогов не наблюдается ни редукции носовых костей черепа, ни соответствующего значительного увеличения носовой вырезки. У носорогов имеется неглубокая низко расположенная, но достаточно обширная предглазничная ямка, служащая для помещения дивертикулюма. По-видимому, хоботок у Lophialetes более развит, чем верхняя губа у Diceros, но функционально слабее, чем у Tapirus. О недостаточной деятельности хоботка свидетельствуют не полностью сросшиеся предчелюстные кости, которая у относительно короче, чем у Tapirus.
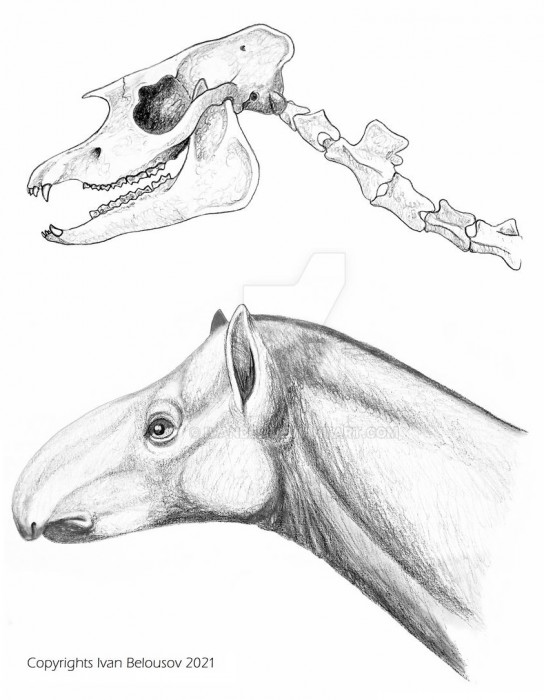
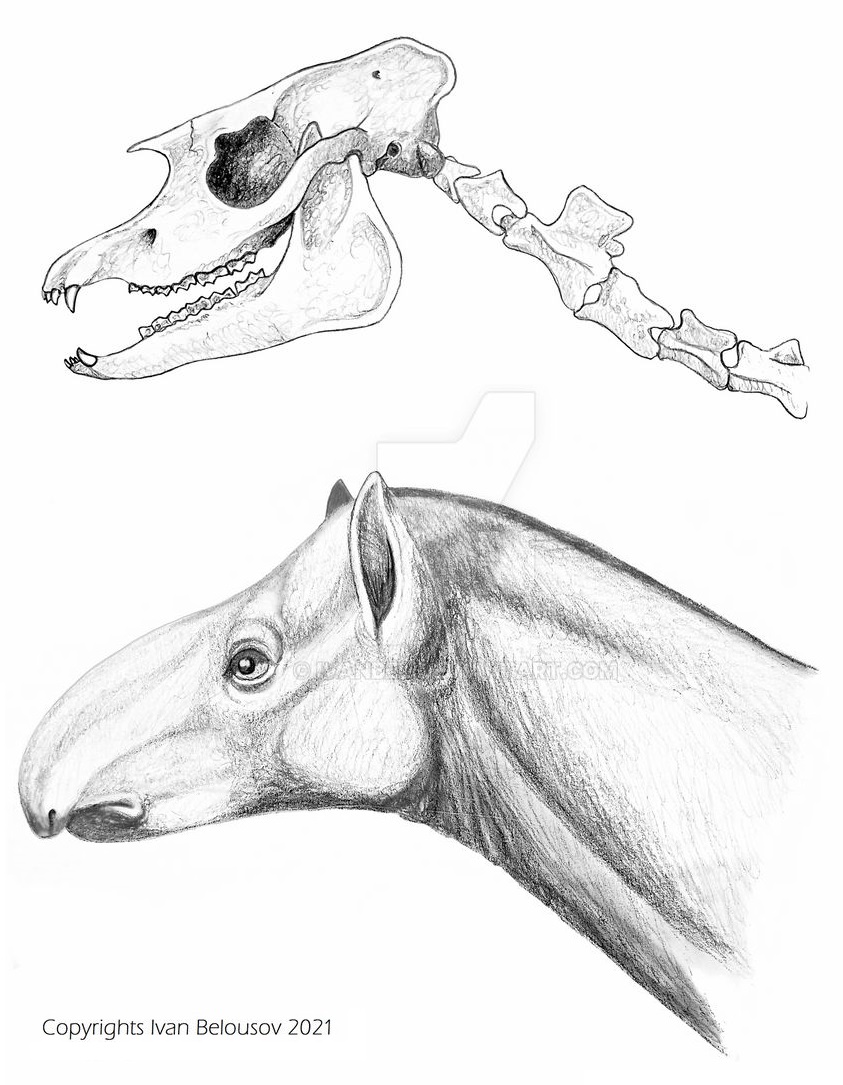
Прорисовка реконструированного черепа с нижней челюстью и шеи, а также портретная реконструкция лофиалета (Lophialetes expeditus). Автор — И. Белоусов.
Последний верхний резец не увеличен, что указывает на его обычную функцию (у Tapirus он бивнеобразный). Носовые кости относительно длиннее, чем у Tapirus. Эти особенности позволяют предположить меньшую механическую нагрузку, которую испытывал лицевой отдел черепа Lophialetes по сравнению с современными тапирами. Помимо этого, лицевой отдел черепа относительно длиннее, чем у Tapirus, а передний край глазницы расположен не над Р4 -М1, а над передним краем М3. Развитие хоботообразной верхней губы у Lophialetes — пример параллелизма внутри надсемейства Tapiroidea. Развитие этого органа происходило независимыми путями у представителей семейства Helaletidae (Helaletes, Colodon) и в более позднее время у настоящих тапиров (Tapiridae). Особенности строения зубной системы Lophialetes и связанные с ними особенности мозговой части черепа и нижней челюсти позволяют судить о способе пережевывания пищи и степени развития челюстной мускулатуры. Движение челюстей в процессе жевания было сложным и включало кругообразные смещения в вертикальной и горизонтальной проекциях.
Посткраниальный скелет
Строение посткраниапьного скелета Lophialetes выделяет его среди эоценовых тапироидов, да и, вероятно, всех ранних непарнопалых. Шейные позвонки (за исключением атланта) абсолютно и относительно длиннее, чем у Heptodon и Hyracotherium. По пропорциям они наиболее близки к шейным позвонкам Deperetella, однако абсолютно и относительно мельче и длиннее последних. Суставные впадины атланта для сочленения с мыщелками черепа глубокие и ориентированы вперед и вверх, в свою очередь черепные мыщелки ориентированы вниз. Сильно развитые затылочные гребни и испещренная шероховатостями бугристая затылочная кость свидетельствуют о прикреплении сильной шейной мускулатуры и связок, причем постановка головы отличалась от таковой у Tapirus. Вероятно, голова была поднята выше и круче наклонена по отношению к оси шеи. На эпистрофее значительно развита передняя суставная поверхность, свидетельствующая о сильном вращении шеей между ним и атлантом.
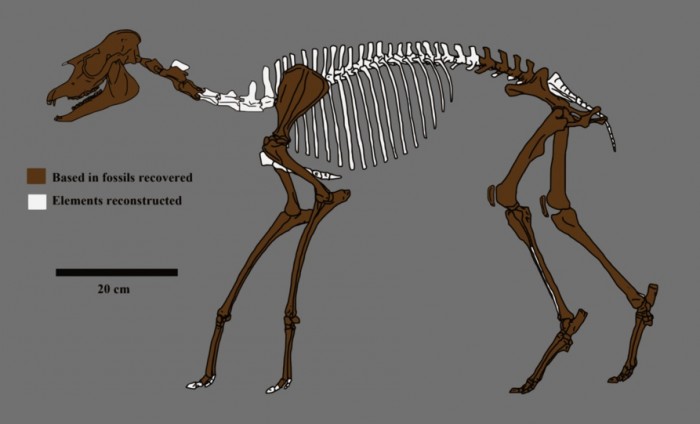
Прорисовка реконструированного скелета лофиалета (Lophialetes expeditus).
Поясничные позвонки Lophialetes представляют переходный тип от предков непарнопалых к более прогрессивным формам. Особенности поясничного отдела способствуют созданию определенной жесткости в этой области позвоночника. Вместе с тем сочленовные отростки еще не образуют настоящего "замка" при соединении позвонков, как у наиболее продвинутых копытных, однако, края передних сочленованных отростков поясничных позвонков подняты вверх в большей степени, чем у Heptodon. Поперечные отростки на поясничных позвонках длинные, ориентированы в горизонтальной плоскости и вперед, но вместе с тем слегка наклонены вниз.
Ограничение изгиба позвоночника в боковую сторону достигается у копытных при помощи разросшихся поперечных отростков, ориентированных горизонтально и вперед. Расширенные окончания поперечных отростков соединены у копытных промежуточной связкой. У хищников и древних предков непарнопалых эти отростки направлены вперед и вниз, а окончания их сужаются. Строение поясничных позвонков Lophialetes показывает значительные преобразования в сторону стабильности поясничного отдела позвоночника при передвижении — умеренный наклон остистых отростков вперед, причем сами отростки расширены, высоко подняты края передних сочленованных отростков, поперечные отростки с расширенными окончаниями и ориентированы вперед и в основном горизонтально. В то же время в строении позвонков поясничного отдела сохраняются черты примитивных предков непарнопалых: сравнительно свободное сочленение позвонков, не образующее "замка" и некоторый наклон поперечных отростков вниз. Эти особенности свидетельствуют о меньшей специализации поясничного отдела у Lophialetes, чем у более поздних непарнопалых (например, лошадей). В настоящее время не представляется возможным определить длину позвоночного столба из-за не полного количества позвонков в коллекциях.
Лопатка
Lophialetes отличается от таковой
Heptodon и
Hyracotherium тем, что обладает более узкой и удлиненной шейкой. Степень развития акромиона свидетельствует о способности отведения передней конечности в сторону. При редукции акромиона боковые движения конечности резко ограничиваются. Например, у лошади (
Equus ferus) акромион отсутствует, так же как и у высших жвачных. Таким образом, у парнопалых и непарнопалых редукция акромиона может свидетельствовать о повышенном значении передне-задних движений конечностью, т. е. нарастающей способности к бегу. Таз
Lophialetes также обладает чертами строения, характерными дня "бегающих" копытных.
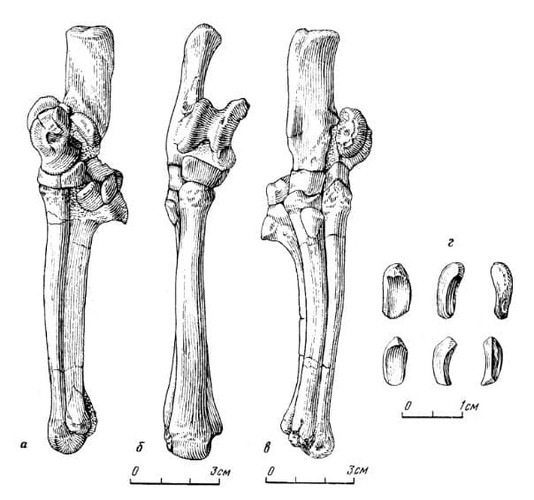
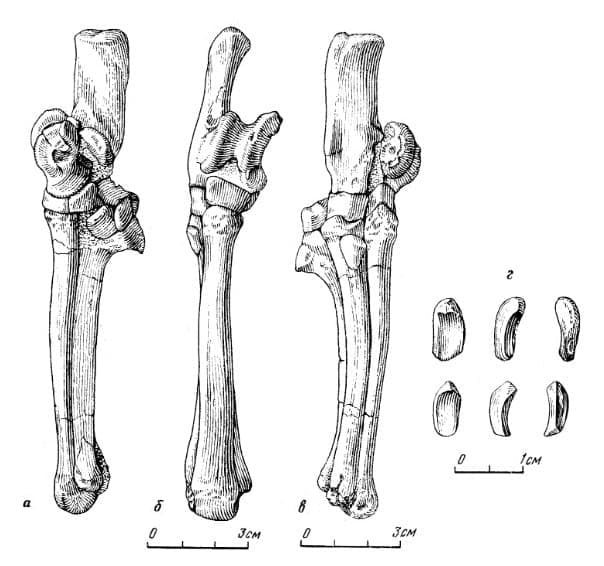
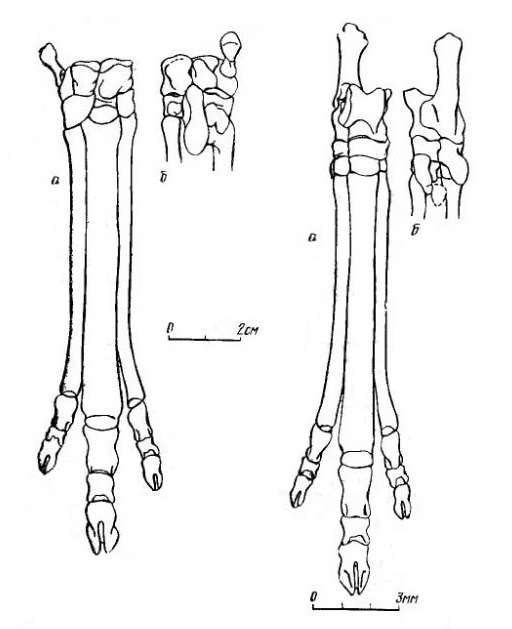
Прорисовки проксимального отдела стопы лофиалета (Lophialetes expeditus) из отложений хайчинской свиты местонахождения Хайчин-Ула III, Монголия: изнутри (а), спереди (б), снаружи (в), сесамовидные кости (г).
Свободные конечности у Lophialetes очень длинные, а отдельные их сегменты тонкие и стройные. Общая длина плеча, лучевой кости и III-й пястной кости в среднем превышает 40 см. Плечевая кость по отношению к лучевой короткая, индекс отношения длины составляет 169,3, что значительно выше, чем у всех известных непарнопалых, а также и парнопалых за исключением современного жирафа (Giraffa camelopardalis) с индексом 160. Однако по абсолютным размерам плечо Lophialetes (11,8 см) приближается к таковым Heptodon (11,5 см) и Нуrаcotherium (97-13 см). Сильно развит проксимальный эпифиз плеча. Суставная поверхность верхнего эпифиза протягивается несколько спереди назад, и конец ее низко опущен назад. Это свидетельствует о значительном сгибании в передне-заднем направлении. Поверхность плеча покрыта шероховатостями и гребнями (местами крепления мышц), которые занимают более проксимальное положение, чем у современного Tapirus terrestris. Их расположение более близко к таковым у некоторых современных жвачных (например, Gazella subgutturosa).
Кисть Lophialetes резко отличается от таковой Hyracotherium, Heptodon и Tapirus. Почти идентична ей кисть Schlosseria, основные отличия которой сводятся к меньшим размерам и пропорциям отдельных костей. Главным отличием в строении кисти от таковых раннеэоценовых форм является практически полная редукция V-й пястной кости, частичное ослабление функции II-й и I-й пястной костей, а вместе с тем усиление среднего пальца и, наконец, общее удлинение пястных костей и фаланг. Довольно высок индекс отношения длины III-й пястной кости к плечевой. У Lophialetes он достигает 73, что значительно превышает отношения у всех известных палеогеновых непарнопалых и приближается к показателю Equus ferus — 78.
Строение посткраниального скелета
Lophialetes убедительно демонстрирует значительную его специализацию, в особенности заметную на конечностях. Как известно, адаптация копытных к быстрому передвижению достигалась путем удлинения дистальных отделов конечности и стабилизации позвоночного столба. При этом происходило упрощение строения кисти и стопы, т. е. сокращение боковых пальцев. По-видимому,
Lophialetes представляет необычно рано для непарнопалых развившийся тип "бегающего" копытного. Серия поясничных позвонков
Lophialetes представляет собой достаточно жесткий на изгиб блок. Строение крыла таза показывает, что бедро сильно сгибалось по отношению к тазу, остававшемуся во время движения относительно иммобильным.
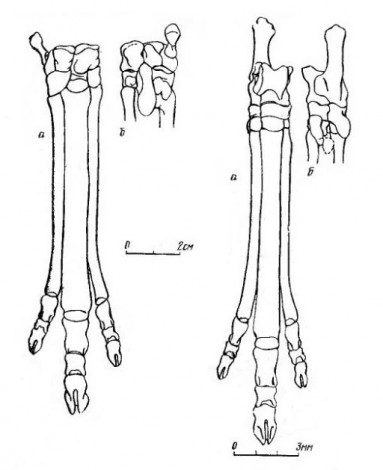
Прорисовки реконструированных правых кисти (слева) и стопы (справа) лофиалета (
Lophialetes expeditus), спереди (а) и сзади (б).
Конечности Lophialetes в целом необычайно длинные и по относительной длине превышают таковые у всех палеогеновых непарнопалых. Тонкие диафизы лучевой и большой берцовой кости сильно изогнуты, чем достигается большая упругость конечности при прыжках (как у современных антилоп). Расстояние между расставленными при шаге конечностями также было необычайно большим. Предположения о значительной способности к бегу подтверждают и некоторая редукция лопаточной ости, сравнительно высокое расположение шероховатости для дельтовидной мышцы на плече, резкое сокращение размеров диафизов локтевой и малой берцовой костей, удлинение пястного и плюсневого отделов кисти и стопы соответственно. Вместе с тем в трехпалых конечностях сильно редуцированы боковые пальцы, слабо развитые суставные поверхности и сильная ассиметрия фаланг которых указывают на относительно малую подвижность этих пальцев. Причем специализация кисти особенно заметна у обсуждаемой формы по сравнению с раннеэоценовыми непарнопалыми, уже обладавшими трехпалой стопой. Эта особенность может быть частично объяснена, если принять во внимание тот факт, что у всех животных "бегающего типа", в том числе у ранних непарнопалых, основная движущая сила обеспечивается задними конечностями, а передние конечности в первую очередь служат как поддержка, на которую опирается тело во время фазы возвращения задних конечностей. По этой причине первоначальная специализация конечностей к бегу у наиболее примитивных непарнопалых, как, например, Hyracotherium была достигнута путем изменения строения стопы, произошедшего, вероятно, еще в позднем палеоцене. Дальнейшую специализацию передних конечностей испытывают более адаптированные к бегу формы (например, Equidae, а среди Ceratomorpha — некоторые Tapiroidea и Hyracodontidae).
Особенностями, противоречащими основной адаптации Lophialetes к бегу, являются отсутствие наружного мыщелка на дистальном блоке плеча, несколько большая по площади надостная ямка (свидетельствующая о сильном развитии надостной мышцы), округлая форма головки бедра. Отсутствие наружного мыщелка, препятствующего у современных "бегунов" боковым движениям конечности, компенсируется у Lophialetes наличием глубокой срединной впадины на дистальном блоке плеча, в которую входит суставной гребень лучевой кости и полулунной вырезки локтевой. Вероятно, это приспособление с не меньшим успехом препятствует боковым сдвигам в локтевом суставе. Относительно большее развитие передней мышцы лопатки у Lophialetes, по-видимому, вызвано необычайным удлинением лучевой кости и связанной с этим необходимостью поднятия конечности. Значительно развиты и спинные отростки шейных позвонков, свидетельствующие о соответствующем развитии шейных мышц. Необычайное удлинение лучевой кости и шейных позвонков — явление, аналогичное таковым у Giraffa и Chalicotheriidae. Однако общая длина свободной передней конечности Lophialetes меньше задней конечности, что отличает его от Giraffa, имеющего обратное соотношение.
Образ жизни
Исходя из особенностей строения скелета Lophialetes, можно предположить, что он обитал в открытых и полуоткрытых пространствах, питаясь наряду с травянистой растительностью побегами кустарников, а также веткам и листьями деревьев, причем часть времени при питании животное могло проводить подобно современной жирафовой газели (Litocranius walleri), опираясь передним конечностями о ствол дерева, либо о нижние его ветви. При этом преимущественное действие получают передние мышцы конечности, причем голова, поднятая высоко вверх, была очень подвижна при захвате пищи с помощью хоботка. Однако в большей степени у Lophialetes выражена адаптация к бегу, что позволяет рассматривать эту форму как экологического предшественника современных антилоп, населявшего эоценовые открытые ландшафты Центральной Азии.
Кормящийся лофиалет (Lophialetes expeditus). Автор — Н. Комаров.
Lophialetes был наиболее подвижным среди других известных тапироидов. Возможно, что он образовывал группы, близкие к территориальному типу, как у современных зебры Греви (Equus grevyi) и дикого осла (Equus asinus), которые не образуют постоянных прочных групп и их состав может меняться в течении нескольких часов, хотя самцы обладают значительным территориальным участком. Морфология лицевого отдела черепа Lophialetes не позволяет предположить наличие у него метящей железы, однако не исключено, что он, подобно современным зебрам и носорогам, использовал другие формы мечения. По-видимому, Lophialetes, наряду с примитивными позднеэоценовыми жвачными — трагулинами (Tragulina), были древнейшими из копытных, которые образовывали стадные ассоциации в условиях зарождавшихся степей в Центральной Азии.
Группа бегущих лофиалетов (Lophialetes expeditus). Автор — Н. Комаров.
Исчезновение
Lophialetes жили во второй половине эоцена в условиях усилившейся аридизации климата Центральной Азии. Отдаленность центральных областей материка от морских побережий позволяет предположить, что в то время климат Центральной Азии приобретал более континентальные черты с большими сезонными колебаниями температуры. По-видимому, тогда начали проявляться более или менее длительные засушливые сезоны. Возможно, что у Lophialetes, как наиболее специализированного рода тапироидов произошли нарушения корреляции организма — развитие центральной нервной системы было еще несовершенно, зубная система также недостаточно специализировалась по сравнению с крайне специализированными (для группы в целом) конечностями.
Подобное несоответствие в строении организма при учете относительно быстрого (по времени) выхода этих тапироидов из "привычных" для их условий обитания — зарослей по берегам рек в открытые пространства могло сыграть роковую роль в их конкурентной борьбе с более совершенными жвачными парнопалыми. Наиболее ранние находки жвачных, представленных трагулинами, в Центральной Азии относятся к среднему и самому началу позднего эоцена. В то время трагулины были еще немногочисленны. Однако во второй половине позднего эоцена они уже широко распространились. Наиболее остро конкурентная борьба между Lophialetes и трагулинами начала проявляться в условиях сезонных изменений и нехватки кормов, когда наступали неурожайные периоды. Пресс хищников в вымирании Lophialetes имел, вероятно, второстепенное значение.
Ссылки
Lophialetes (Paleobiology Database)
перейти
Литература
Зоогеография палеогена Азии / Отв. ред. Крамаренко Н. Н. — М.: Наука, 1974
Основы палеонтологии. В 15 т. Т. 13: Млекопитающие / Гл. ред. Орлов Ю. А. — М.: Государственное научно-техническое издательство литературы по геологии и охране недр, 1962
Решетов В. Ю. Раннетретичные тапирообразные Монголии и СССР. — М.: Наука, 1979
Radinsky L. B. Early tertiary Tapiroidea of Asia. Bulletin of the American museum of Natural History 129, 1965