Макраухении — Macrauchenia
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Клада Скротиферы (Scrotifera)
Клада Феревунгуляты (Fereuungulata)
Клада Эвунгуляты (Euungulata)
Клада Мезаксонии (Mesaxonia)
Клада †Южноамериканские копытные (Meridiungulata)
Отряд †Литоптерны (Litopterna)
Надсемейство †Макраухениоиды (Macrauchenioidea)
Семейство †Макраухениевые (Macraucheniidae)
Подсемейство †Макраухениины (Macraucheniinae)
Род †Макраухении (Macrauchenia)
Вид †Macrauchenia patachonica
Вид †Macrauchenia ullomensis
Макраухении (Macrauchenia Owen, 1838) — род подсемейства макраухениин (Macraucheniinae Gervais, 1855), происходящий позднего миоцена - раннего голоцена Южной Америки. Надежно установлены 2 вида.

Реконструкция внешнего облика макраухении (Macrauchenia patachonica). Автор — Р. Евсеев.
История изучения
Остатки макраухении впервые были обнаружены 9 февраля 1834 г. в порту Сан-Хулиан в Патагонии (территория современной Аргентины). Молодой натуралист Ч. Дарвин участвовал в кругосветной экспедиции на корабле «Бигль» под управлением капитана Р. Фицроя. Пока «Бигль» выполнял гидрографическую съемку, Дарвин исследовал отложения Луханской формации недалеко от гавани и нашел в них несколько шейных и спинных позвонков, а также элементов задней конечности. Не являясь специалистом, он предварительно определил находку как "какое-то крупное животное, мне кажется, мастодонт". В будущем эта находка стала одним из открытий, на основе которых Дарвин со временем разработал свою эволюционную теорию.
В 1837 г., вскоре после возвращения «Бигля» в Англию, анатом Р. Оуэн установил, что кости на самом деле принадлежат млекопитающему, напомиеающему внешне ламу или верблюда, которого этот ученый назвал Macrauchenia patachonica. Родовое наименование образовано древнегреческими словами µακρος (makros) — "большой, длинный", и αυχην (auchèn) — "шея", означая таким образом "длинная шея". Кроме того, auchenia — этот старое латинское название для южноамериканских верблюдовых (ламы, викуньи, альпаки). Видовой эпитет указывает на место находки окаменелостей.
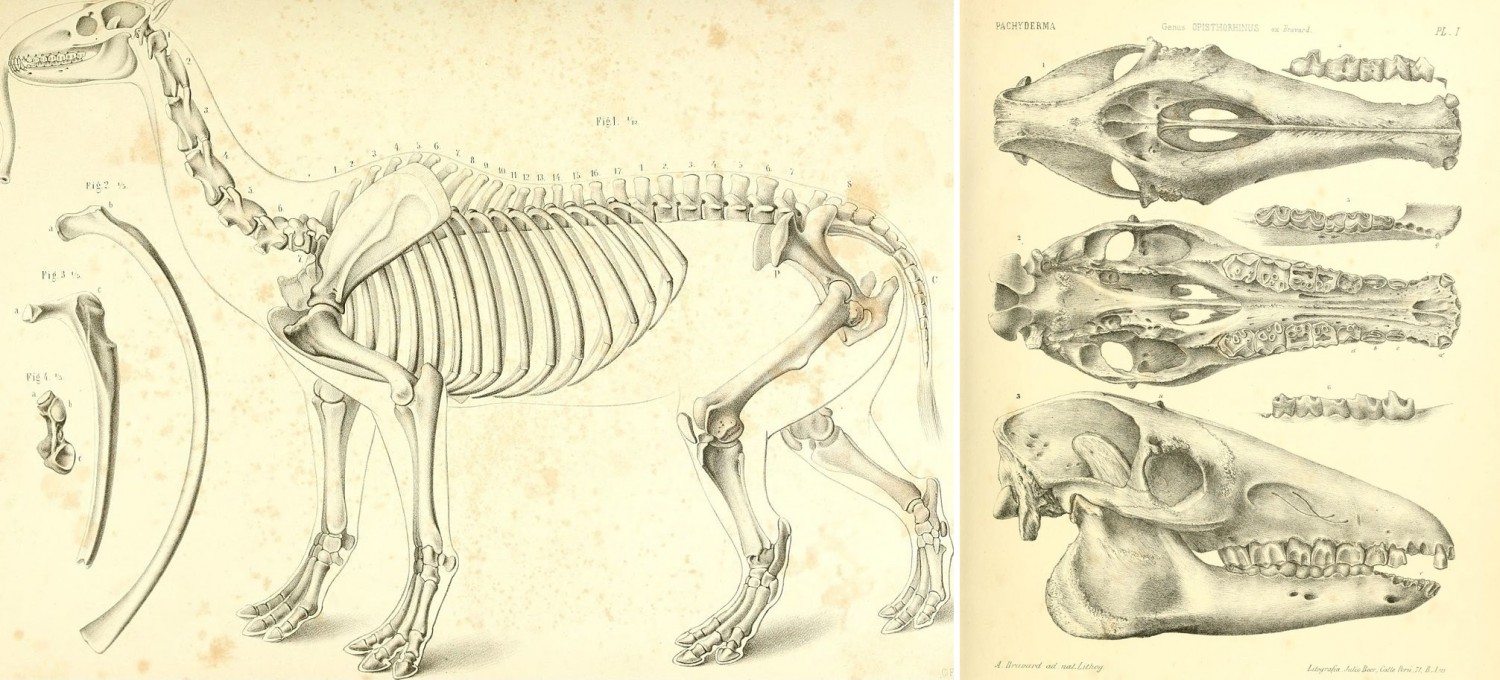
Старинные прорисовки скелетных остатков макраухении (Macrauchenia patachonica).
В дальнейшем преимущественно из Патагонии, а также ряда других районов Южной Америки, было описано еще несколько видов рода Macrauchenia, большинство которых со временем признали недействительными либо реклассифицировали. От типового вида собраны полные скелеты, тогда как остатки других представителей рода довольно фрагментарны.
Строение и функции
Размеры макраухений крупнейшие для отряда. Длина их черепа составляет 45-55 см, общая длина тела достигала 3,2 м, а высота в холке — 1,8 м. Масса оценивается в 500-1000 кг. Внешне этих копытные несколько походили на современных верблюдов (Camelus), но отличались от них большей массивностью.



Смонтированный скелет макраухении (Macrauchenia patachonica).
Череп относительно тела небольшой. Его лицевой и мозговой отделы вытянуты, особенно длинная морда. Носовые кости очень маленькие, рудиментарные, а носовые отверстия сдвинуты далеко назад и образуют большие дыры, расположенные в верхней части черепа между глазницами (сдвинутые вверх ноздри — характерная эволюционная тенденция поздних Macraucheniidae). Глазницы замкнуты. Челюсти несколько расширены в своей передней части.

Череп с нижней челюстью макраухении (Macrauchenia patachonica). Длина его составляет 53 см; большая часть передних зубов отсутствует.
Зубная формула: I3/i3, C1/c1, P4/p4, M3/m3 = 44. Коронки зубов относительно невысокие, но выше, чем у других литоптерн. Зубы сидят плотно друг к другу, и лишь между последним верхним резцом и клыком находится небольшая диастема. На внешней стенке хорошо развитых щечных зубов расположены две глубоких лунки, от которых идут, направляясь назад, стертые гребни, образующие при стирании замкнутые кружочки. Нижние щечные зубы также содержат по две лунки.
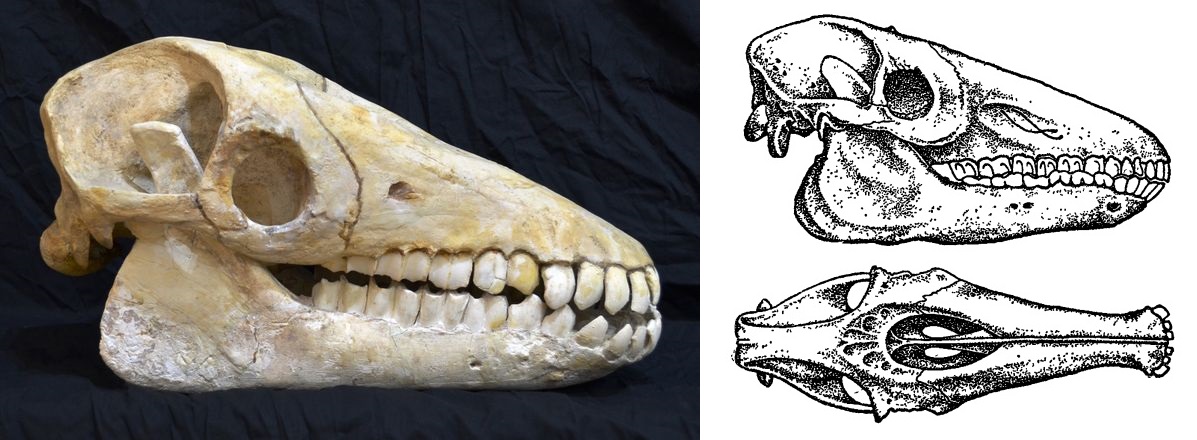
Череп с нижней челюстью макраухении (Macrauchenia patachonica) и его прорисовки, сбоку и сверху.
Шея длинная, ее позвонки имеют плоские сочленовные поверхности. Грудные позвонки обладают достаточно высокими остистыми отростками. Конечности длинные и мощные. Их передняя и задняя пары имеют примерно одинаковую длину. При этом передняя нога более короткая в своем верхнем отделе и длинная в нижнем, что указывает на приспособленность к бегу. Однако задняя нога значительно более длинная в верхнем и пропорционально короткая в нижнем отделах, в результате чего задние ноги не подходят для быстрого бега. Существенная разница в строении между передними и задними конечностями позволяет предполагать, что макраухении более полагались не на скорость бега, а на его маневренность. Это было особенно полезно, если эти копытные населяли районы со сложным рельефом местности, что является опасным для быстроходных животных, которые не могли столь же устойчиво держаться на ногах.
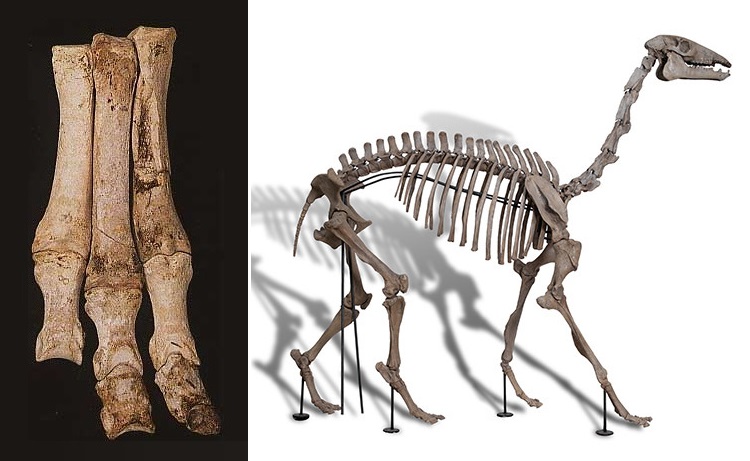
Слева: кости нижней части конечности макраухении (Macrauchenia patachonica), найденные Ч. Дарвином на территории Аргентины во время путешествия на корабле «Бигль». Общая длина этого образца составляет 30,5 см.
Справа: смонтированный скелет макраухении (Macrauchenia patachonica).
Строение костей голени и значительная подвижность голеностопного сустава дополнительно может свидетельствовать о способности резко поворачиваться, тем самым быстро меняя направление передвижения. Карпальные кости не смещены относительно друг друга. Кисть и стопа несут по три хорошо развитых пальца, из которых наибольшую длину имеет средний. Пальцы касались земли концевыми фалангами, несущими копыта. Судя по строению, напоминающему таковое у ныне живущих носорогов, ступни не особенно хорошо приспособлены к бегу, а больше подходят для поддержания значительной массы тела.

Вверху: прорисовка реконструированного скелета и телесных очертаний макраухении (Macrauchenia patachonica). Автор — И. Иофрида.
Внизу: реконструкция внешнего облика макраухении (Macrauchenia patachonica). Автор — Р. Евсеев.
Цветовая окраска макраухений неизвестна. Предположения об этом можно строить лишь на основании ныне существующих копытных, которые обитают в аналогичных условиях, таким как ламы в Южной Америке и антилопы в Африке. На основании этого полагают, что макраухении имели песочный окрас.
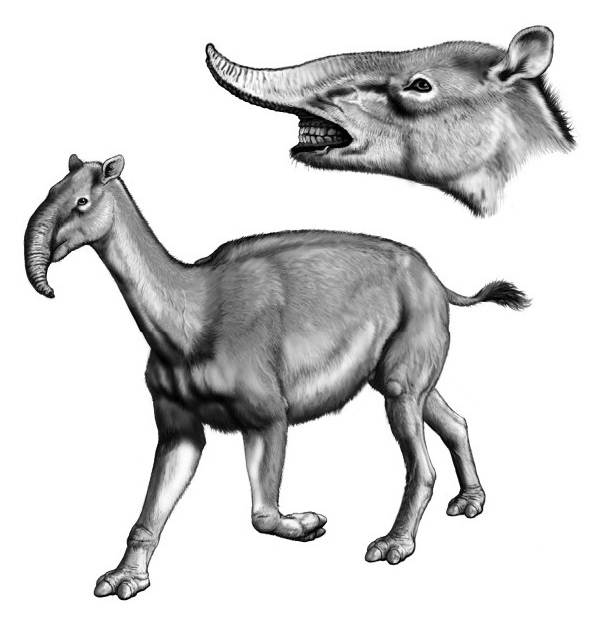
Реконструкция внешнего облика и портретная реконструкция макраухении (Macrauchenia patachonica). Автор — К. Бьюэлл.
Состав
Macrauchenia patachonica Owen, 1838 (тип) — типовой вид, известный из позднего плиоцена - позднего плейстоцена Аргентины (провинции Санта-Крус, Кордова, Энтре-Риос, Буэнос-Айрес), Боливии (департаменты Ла-Пас, Тариха, Чукисака), Парагвая (департамент Пресиденте-Аес), Бразилии (штаты Пернамбуку, Пиауи, Риу-Гранди-ду-Сул), Уругвая (департаменты Сальто, Монтевидео), Перу (регион Куско) и Чили. Синоним: Macrauchenia patagonica Lydekker, 1894.

Смонтированный скелет макраухении (Macrauchenia patachonica) (образец AMNH 14489); на переднем плане — смонтированный скелет примитивного копытного фенакода (Phenacodus primaevus), а также черепа с нижними челюстями литоптернов теозодона (Theosodon patagonicum) и тоатерия (Thoatherium crepidatum). Американский музей естествознания, Нью-Йорк.
Macrauchenia ullomensis Sefve, 1914 — вид, известный из плейстоцена (включая ранний) Боливии (департаменты Ла-Пас, Оруро).
Остатки, определенные лишь до рода и обозначенные Macrauchenia spp., известны из позднего миоцена / плиоцена (монтеэрмосский / чападмалальский век) - раннего голоцена Боливии (департаменты Оруро, Ла-Пас, Чукисака), Аргентины (провинции Жужуй, Буэнос-Айрес, Кордова, Сантьяго-дель-Эстеро), Бразилии (штаты Пернамбуку, Параиба, Пиауи), Перу (регион Пьюра), Уругвая, Венесуэлы (штат Фалькон) и Чили (область Магальянес).

Смонтированный неполный скелет макраухении (Macrauchenia patachonica).
Macrauchenia antiqua Ameghino, 1887 — сомнительный вид, описанный из плейстоцена Аргентины.
Macrauchenia formosa Moreno, 1888 — сомнительный вид, описанный из плейстоцена Аргентины.
Macrauchenia intermedia Moreno, 1888 — сомнительный вид, описанный из плейстоцена Аргентины.
Macrauchenia antiqua Ameghino, 1887 — вид, описанный из плейстоцена Аргентины. Синоним: Macrauchenia antiquus. Выделен в самостоятельный род Promacrauchenia Ameghino, 1904.
Macrauchenia boliviensis Huxley, 1860 — вид, описанный из плейстоцена Боливии. В настоящее время считается таксоном nomen dubium.

Области распространения остатков макраухениин Macrauchenia (красный цвет) и Xenorhinotherium (желтый цвет).
Образ жизни
Судя по географии находок, макраухении были широко распространены, встречаясь от юга Чили до северо-востока Бразилии и побережья Венесуэлы. Эти животные населяли саванноподобные ландшафты от сухих до влажных, но более всего тяготели к пампасам на юго-востоке Южноамериканского континента. В Боливии ископаемые остатки Macrauchenia ullomensis найдены на высоте более 4000 м над уровнем моря, что предполагает переносимость достаточно низких температур. Обнаружение расположенных в непосредственной близости остатков 5 молодых особей в одном из местонахождений на севере Чили может указывать на их жизнь стадами или семейными группами.
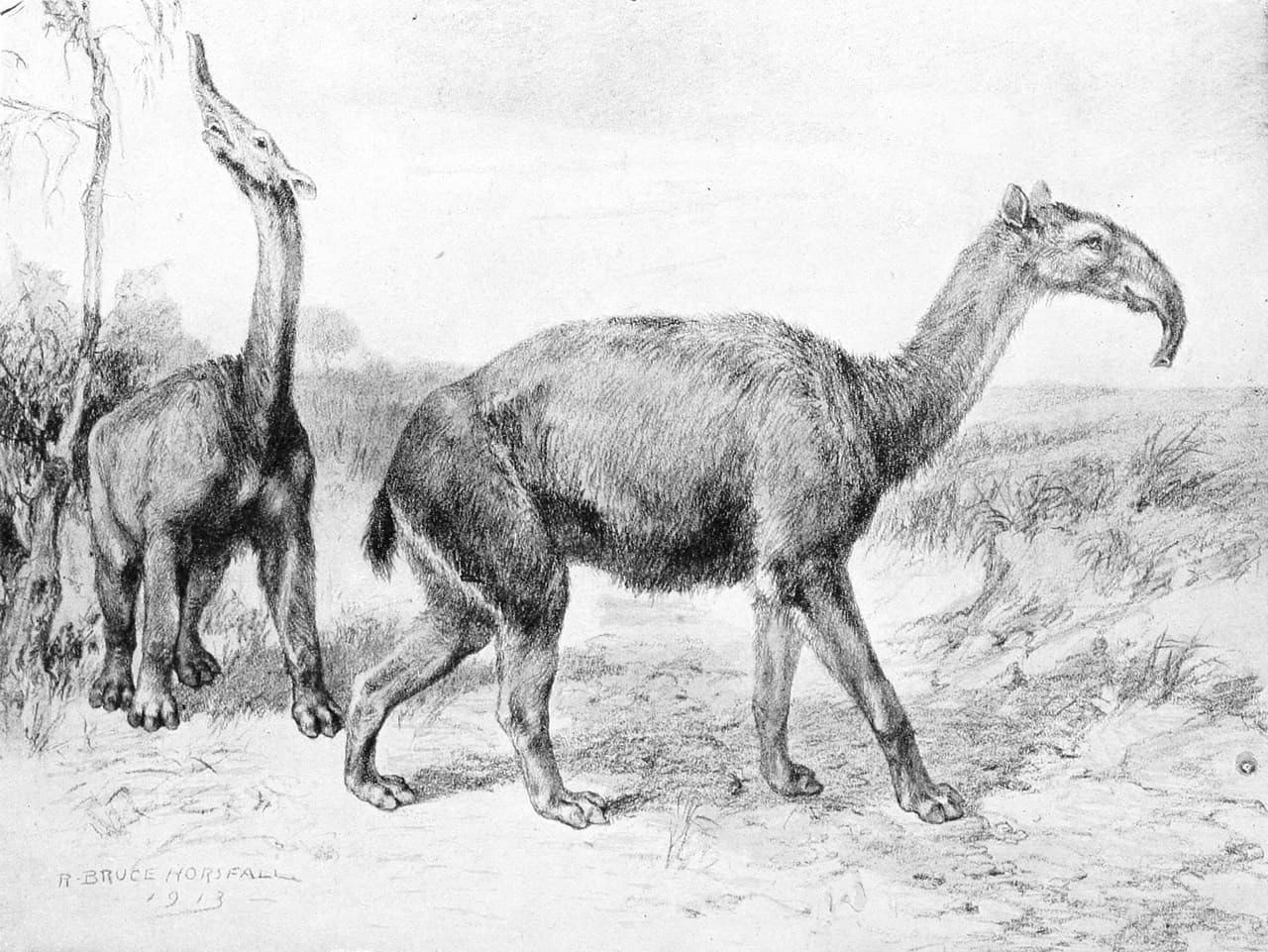
Макраухении (Macrauchenia patachonica) в естественном окуржении. Автор — Р. Хорсфалл, 1913 г.
Раньше некоторые исследователи полагали, что макраухении вели полуводный образ жизни, являясь обитателями болот. По их мнению, ноздри этих животных располагались наверху головы для дыхания в то время, когда тело было погружено в воду, а между пальцами были натянуты плавательные перепонки. Однако в дальнейшем эта гипотеза была отвергнута. У ныне живущих копытных длинная шея является адаптацией, позволяющей дотягиваться до веток деревьев и кустарников, а также издали замечать потенциальную опасность. Судя по всему, сходное поведение было характерно и для макраухений.
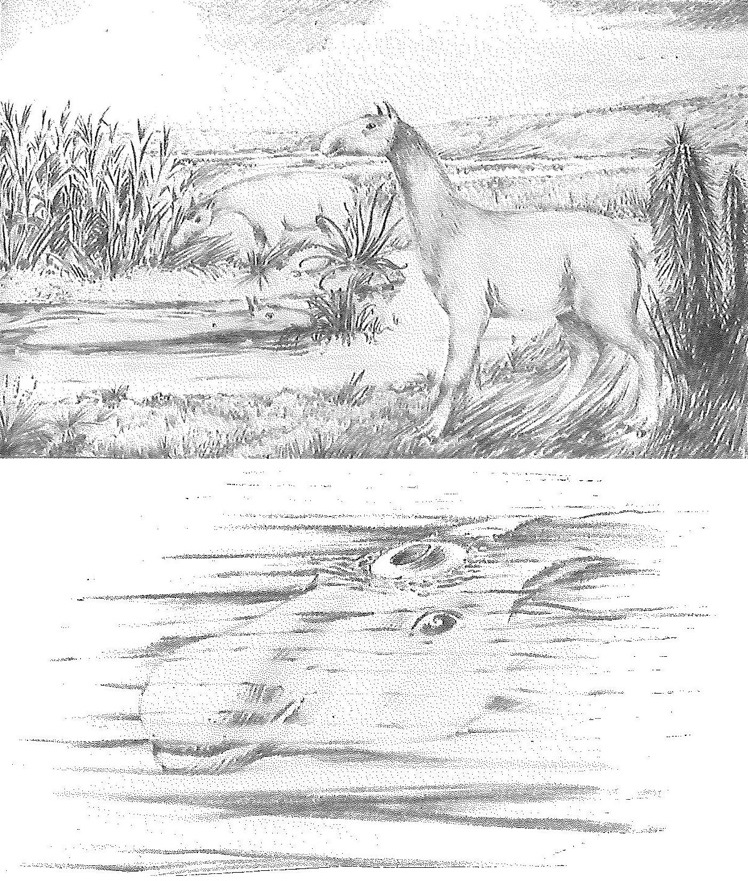
Реконструкции, отображающие воззрения о полуводном образе жизни макраухении (Macrauchenia patachonica).
Возможно, высоко расположенные ноздри указывают на наличие небольшого хоботка, напоминающего таковой современных тапиров (Tapirus spp.) или сайгака (Saiga tatarica), также обладающих сдвинутыми назад носовыми отверстиями. Полости в лобных костях непосредственно позади носовых отверстий макраухений могут являться местами прикрепления хорошо развитых мышц, поддерживающих хоботок. Он мог использоваться для захвата листвы и прочей растительности, либо препятствовал попаданию в легкие пыли. Вместе с тем, спереди морда макраухений образована сплошной костью, а их удлиненная шея позволяла тянуться вверх, чего не наблюдаются ни у одного ныне живущего млекопитающего с хоботом.

Макраухении (Macrauchenia patachonica) в естественном окружении. Автор — Р. Евсеев.
Альтернативная точка зрения состоит в том, что макраухении и их родственники были лишены хоботка, напоминая очертаниями морды современных лосей (Alces spp.) или верблюдов (Camelus spp.). Они могли кормиться, объедая жесткую и колючую растительность, а поднятые на макушку головы ноздри позволяли им доставать листья, не повреждая нос. Динозавры-завроподы, реконструируемые как поедатели хвойных и саговников, имеют похожие морды с высоко расположенными ноздрями, а современные жирафы (Giraffa spp.) и антилопа геренук (Litocranius walleri), объедающию колючую растительность, обладают более втянутыми внутрь морды ноздрями, чем родственные им таксоны с другим характером питания.


Слева: следовой отпечаток макраухении (Macrauchenia sp.).
Справа: семейство макраухений (Macrauchenia patachonica) на водопое.
Форма морды, устройство зубного аппарата, особенности микроизноса и изотопный состав эмали, а также низкий уровень гипсодонтии зубов свидетельствует, что рацион Macrauchenia patachonica включал как листья деревьев и кустарников (С3-растительность), так и различные травянистые растения (С4-растительность).

Группа макраухений (Macrauchenia patachonica), вариант реконструкции с далеко сдвинутыми ноздрями. Автор — В. Симеоновски.
Из числа плотоядных макраухении первоначально соседствовали с форораками и спарассодонтами. Тогда как первые из них были хорошими бегунами, вторые являлись засадными охотниками, вероятно, неспособными догнать добычу, если она сумела уклонилась от их первой атаки. После формирования Панамского перешейка в конце плиоцена - начале плейстоцена на Южноамериканском континенте переселяются медведи, собаки и кошки. По-видимому, от хищников эти копытные спасались посредством маневренного бега, а при невозможности ретироваться отбивались с помощью сильных конечностей. Кроме того, крупный размер взрослых особей уже сам по себе ограничивал их уязвимость для большинства хищников. Молодые животные являлись гораздо более уязвимыми. Так или иначе, макраухении смогли ужиться с новыми, более прогрессивными формами плотоядных и существовали бок о бок с ними на протяжении нескольких миллионов лет. Последнее в равной степени справедливо и в отношении возможных конкурентов из числа пришедших с севера копытных — например, верблюдов и лошадей. Так, остатки макраухений часто находят в непосредственной близости с таковыми одного родов верблюжьих, Palaeolama.

Макраухении (Macrauchenia patachonica) спасаются бегством от охотящихся смилодонов (Smilodon populator). Автор — М. Антон.
Вымирание
Макраухении исчезли примерно на границе плейстоцена с голоценом, являясь таким образом последними достоверно известными представителями литоптерн. Их наиболее молодые остатки, найденные на юге Чили, имеют возраст около 11,7 тыс. лет. По времени исчезновение этих животных совпало с вымиранием большей части южноамериканской мегафауны. Оно связывается главным образом с глобальными климатическими изменениями. Определенную роль могла сыграть и охотничья деятельность проникшего в Южную Америку около 15 тыс. лет назад первобытного человека, в очагах которого иногда находят обожженные кости макраухений.
Генетика
В 2017 г. команде исследователей удалось секвенировать из костей макраухении, найденной в пещере Баньо-Нуэво 1 на юге Чили, около 80 % ее митохондриального генома. Выяснилось, что это животное (как и остальные литоптерны) относится к сестринской по отношению к непарнопалым группе, с предполагаемой датой расхождения их эволюционных линий в 66 млн. лет назад. Ранее к аналогичному выводу пришли ученые, применившие методы палеопротеомики и помимо литоптерн включившие в сестринскую к непарнопалым кладу нотоунгулят. Проведенный ими анализ последовательностей белка соединительной ткани коллагена показал, что ближайшими родственниками Macrauchenia patachonica из литоптерн и Toxodon platensis из нотоунгулят являются лошади и носороги, но время разделения их эволюционных линий было отнесено к эоцену (около 55 млн. лет назад).
Ссылки
Макраухении (Википедия) перейти
Macrauchenia (Paleobiology Database) перейти
Литература
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 3 — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 13: Млекопитающие / Гл. ред. Орлов Ю. А. — М.: Государственное научно-техническое издательство литературы по геологии и охране недр, 1962
Павлова М. В. Палеозоология. Ч. 2. Позвоночные. — М. Л.: Государственное издательство, 1929
Ромер А. Ш. Палеонтология позвоночных. — М. — Л.: Государственное научно-техническое издательство нефтяной и горно-топливной литературы, 1939
Симпсон Дж. Великолепная изоляция. История млекопитающих Южной Америки. — М.: Мир, 1983
Тернер А., Антон М. Большая энциклопедия доисторических животных. — М.: Оникс, 2006