Мегаладаписы — Megaladapis
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Суперотряд Эвархонтоглиресы (Euarchontoglires)
Грандотряд Эвархонты (Euarchonta)
Миротряд Приматообразные (Primatomorpha)
Отряд Приматы (Primates)
Подотряд Мокроносые приматы (Strepsirrhini)
Инфраотряд Лемурообразные (Lemuriformes)
Надсемейство Лемуроиды (Lemurоidеа)
Семейство †Мегаладаповые (Megaladapidae)
Род †Мегаладаписы (Megaladapis)
Подрод †Пелориадаписы (Peloriadapis)
Вид †Мегаладапис Эдвардса (Megaladapis edwardsi)
Подрод †Мегаладаписы (Megaladapis)
Вид †Мадагаскарский мегаладапис (Megaladapis madagascariensis)
Вид †Мегаладапис Грандидье (Megaladapis grandidieri)
Мегаладаписы (Megaladapis Forsyth-Major, 1894) — род монотипического семейства мегаладаповых (Megaladapidae Forsyth-Major, 1894), происходящий из позднего плейстоцена - голоцена Мадагаскара. Установлены 2 подрода с 3 видами.

Реконструкция внешнего облика мегаладаписа Эдвардса (Megaladapis edwardsi). Автор — Р. Евсеев.
Синонимы: Peloriadapis Grandidier, 1899; Paleolemur Lorenz von Liburnau, 1900; Mesoadapis Lorenz von Liburnau, 1900; Megalindris Standing, 1908.
История изучения
Впервые череп мегаладаписа был описан зоологом Ч.И. Форсайтом-Мейджором в докладе Лондонскому королевскому обществу в июне 1893 г., а затем в журнале «Philosophical Transactions of the Royal Society B» за 1894 г. Череп был получен Британским музеем в составе коллекции фоссилий с Мадагаскара. Дж. Т. Ласт, собиравший эту коллекцию для финансового магната У. Ротшильда, сообщил, что череп был найден в болотах Амбулисатра близ юго-западного побережья острова. Предложенное Форсайтом родовое имя Megaladapis (от древнегреческого μέγας — "большой" — и Adapis) было связано с тем, что новый таксон обладал рядом общих характеристик (в том числе в строении коренных) с описанным ранее родом ископаемых приматов Adapis. Новоописанному виду Форсайт дал название Megaladapis madagascariensis. Уже в первой публикации Форсайта отмечались многочисленные общие черты в строении черепа нового таксона и современного коалы (Phascolarctos cinereus) (хотя у мегаладаписа эти признаки сильно гипертрофированы), а также вероятное сходство в способе их передвижения и составе рациона, из-за чего мегаладаписов в дальнейшем также стали называть коаловыми лемурами.

Череп мадагаскарского мегаладаписа (Megaladapis madagascariensis).
В статье 1894 г. Форсайт подчеркивал, что данное им название не должно трактоваться как указывающее на близкую родственную связь мегаладаписов с адаписами; напротив, он отчетливо понимал, что эти таксоны далеки друг от друга, хотя и считал, что адаписы представляют собой ветвь лемурообразных. Форсайт отмечал сходство по отдельным морфологическим характеристикам между мегаладаписами и такими современными представителями лемурообразных, как лепилемуровые (Lepilemuridae), карликовые лемуры (Cheirogaleidae) и индриевые (Indriidae). Тем не менее он выделял их в общей классификации лемуров в отдельное семейство — мегаладаповые (Megaladapidae).

Неполный смонтированный скелет мегаладаписа Эдвардса (Megaladapis edwardsi).
С родом Megaladapis с начала XX в. и вплоть до 1970-х гг. связывали в качестве младшего синонима также описанный в 1895 г. А. Фильолем по неполной плечевой кости вид Thaumastolemur grandidieri, но позже была доказана ошибочность этой точки зрения. В 1898 г. французский натуралист А. Грандидье описал по нескольким зубам и фрагменту челюсти вид Peloriadapis edwardsi, который затем был связан с родом Megaladapis. Название Peloriadapis сохраняется как подродовое для самого крупного известного вида мегаладаписов. В следующие несколько лет были описаны новые виды мегаладаписов, а также несколько таксонов родового ранга, в дальнейшем стали рассматриваться в качестве младших синонимов рода Megaladapis. В 1902 г. Грандидье провел объединительную работу, в частности объединив описанный Форсайтом вид Megaladapis insignis со своим. В 1903 г. был установлен третий вид мегаладаписов, считающийся валидным и сегодня — Megaladapis grandidieri. Г. Стэндинг описал его по находкам, сделанным в Ампасамбазимбе в центральной части острова.

Смонтированный скелет мегаладаписа Грандидье (Megaladapis grandidieri).
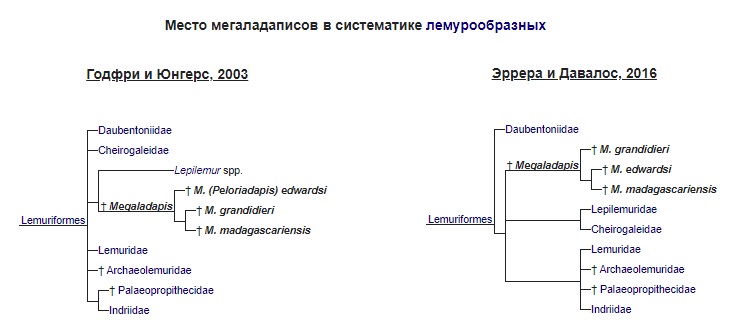
Родственные связи
Долгое время мегаладаписы рассматривались в качестве сестринской группы по отношению к тонкотелым лемурам (Lepilemur) и объединялась с ними в одно семейство, которое могли именовать как Lepilemuridae, так и Megaladapidae, а также выделять внутри него 2 подсемейства — Lepilemurinae и Megaladapinae соответственно. Однако исследования конца XX - начала XXI вв., использовавшие методы анализа палео-ДНК и молекулярные часы, этот постулат не подтвердили. Согласно этим исследованиям мегаладаписы являются сестринским таксоном либо для лемуровых (Lemuridae), либо даже для всех групп лемурообразных (Lemuriformes) после отделения руконожковых (Daubentoniidae) от основного филогенетического ствола.
Строение и функции
Размеры мегаладаписов очень крупные для мокроносых приматов: в зависимости от вида, длина черепа составляет 23-32 см, общая длина варьирует в пределах 100-150 см, а вес оценивается в 40-85 кг. До того, как были разрешены проблемы с принадлежностью костей скелета, большие черепа являлись причиной того, что общие размеры тела и масса оценивались как еще более значительные. Мегаладаписы имели приземистое тело и крепкое, плотное сложение. Морфологически они, как и другие крупные субфоссильные лемуры, значительно больше походили на более мелких современных лемуров, чем на сравнимых с ними по величине человекообразных обезьян, таких как орангутаны (Pongo) и гориллы (Gorilla).

Смонтированные скелеты мегаладаписа Эдвардса (Megaladapis edwardsi).
Череп, непропорционально большой по отношению к телу, непохож по строению с таковым других лемуров и скорее напоминает череп копытных. Лицевой отдел сильно удлинен, мозговой — сравнительно небольшой. Череп несколько изогнут в верхне-нижнем направлении и значительно сжат с боков. Носовые кости приподняты и загнуты вниз, далеко выдаваясь за переднюю границу твердого неба. Такая форма носовых костей уникальна для приматов и имеет сходство с анатомией свиного рыла или морды носорога (но не хоботка тапиров). Подглазничное отверстие невелико. Глазницы подняты высоко вверх и отстоят далеко друг от друга. Они направлены в стороны, так что бинокулярность зрения почти не выражена. Скуловые дуги массивные. Имеются хорошо развитые сагиттальный и затылочный гребни. Слуховые барабаны уплощены. Имеется костный канал, соединяющий барабанную полость с внешней средой.

Череп с нижней челюстью мегаладаписа Эдвардса (Megaladapis edwardsi).
По-видимому, височная мышца в своей задней части была очень мощной, тогда как в передней части несколько редуцирована. Длинный, сильно выступающий отросток затылочной кости указывает на увеличенные размеры двубрюшной мышцы — явления, распространенного среди средних и крупных лемуров в целом, но у мегаладаписов достигающего своего пика. Засуставный отросток скуловой дуги также необычайно мощный, с широкой передней поверхностью. Большое затылочное отверстие сдвинуто назад, затылочные мыщелки расположены перпендикулярно основанию черепа. Нижняя челюсть относительно массивнее, чем у горилл (Gorilla), с глубоким телом и сильно развитой восходящей ветвью. Подбородочный симфиз у взрослых особей сросшийся. Сочленовный отросток по форме напоминает таковой ныне живущих тонкотелых лемуров (Lepilemur).
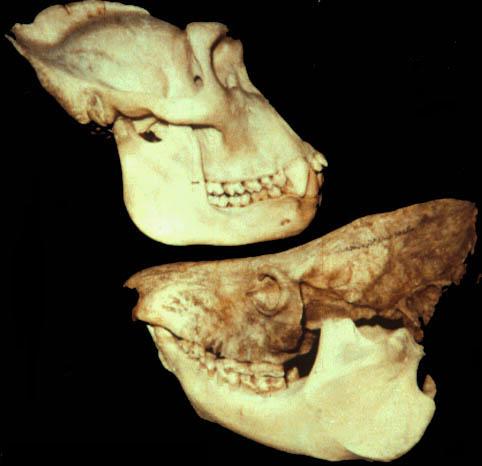
Черепа с нижними челюстями западной гориллы (Gorilla gorilla) (вверху) и мегаладаписа Эдвардса (Megaladapis edwardsi) (внизу) в сравнении.
Зубная формула: I0/i2, C1/c1, P3/р3, M3/m3. Верхние резцы отсутствуют; возможно, вместо них на передней части верхней челюсти имелись режущие роговые пластины, как у некоторых жвачных. Верхний клык большой, загнутый назад, с острой вершиной и режущей кромкой. Нижние резцы длинные и узкие, образуют почти горизонтальный "гребешок". Нижний клык крупный, хотя и уступает по размерам верхнему. Между клыками и щечными зубами в обеих челюстях расположены широкие диастемы. Предкоренные крупные и остроконечные, особенно клыковидный р1. Коренные огромных размеров со сложными бугорками и хорошо выраженными гребнями. Наблюдается существенное увеличение размеров щечных зубов от передних к задним.

Этапы портретной реконструкции мегаладаписа Эдвардса (Megaladapis edwardsi). Автор — В. Симеоновски.
Сдвинутое к задней стенке черепа затылочное отверстие по сути превращает вытянутую голову мегаладаписа в продолжение длинной шеи. Туловище удлинено. Таз своими очертаниями несколько напоминает таковой копытных. Конечности массивные, но сравнительно короткие в сравнении с вытянутым туловищем — в особенности это касается плечевых костей, сильно укороченных в сравнении с аналогичными костями у современных обезьян сопоставимых размеров, однако и бедренная кость у мегаладаписов короче относительно размеров тела. Передние конечности длиннее задних: интермембральный индекс (соотношение длин передней и задней конечностей) варьирует в пределах 114-120. Круральный индекс (соотношение длин большеберцовой и бедренной костей) составляет 73-90. Все конечности слегка выгнутые, что характерно для лазающих животных. Длина кистей и стоп относительно общей длины конечностей непропорционально велика. Пальцы крепкие, с удлиненными и искривленными фалангами. I-е пальцы как на кисти, так и на стопе длинные и сильные, "клешневидные" — значительно противопоставлены остальным. Хвост сильно укорочен. Половой диморфизм по размерам тела или клыков почти не выражен.
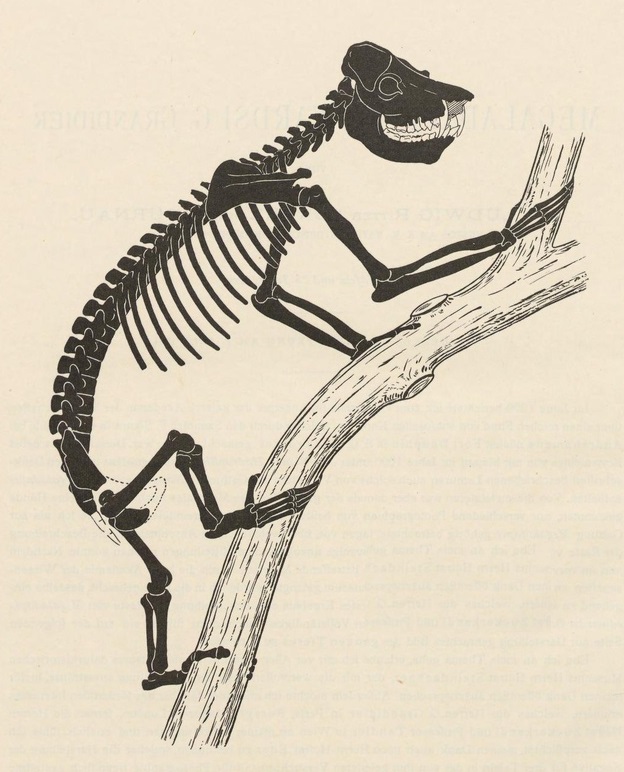
Прорисовка частично реконструированного скелета мегаладаписа Эдвардса (Megaladapis edwardsi).
Состав
В настоящее время валидными считаются 3 вида мегаладаписов, которых нередко разносят в 2 подрода — пелориадаписов (Peloriadapis Grandidier, 1899) с видом Megaladapis edwardsi и собственно мегаладаписов (Megaladapis Forsyth-Major, 1894) с видами Megaladapis madagascariensis и Megaladapis grandidieri.
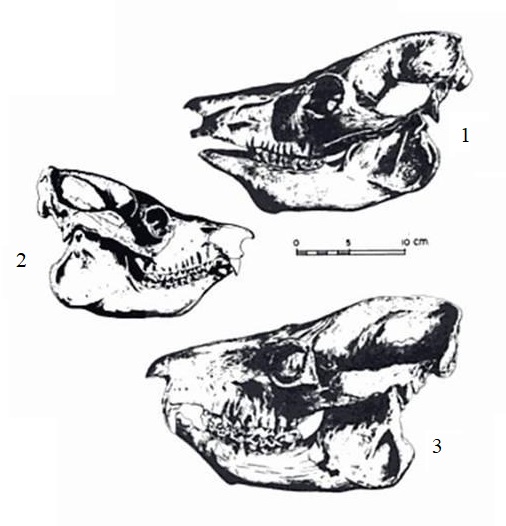
Прорисовки черепов с нижними челюстями различных видов мегаладаписов, приведенных в общем масштабе: 1 — мегаладапис Грандидье (Megaladapis grandidieri); 2 — мадагаскарский мегаладапис (Megaladapis madagascariensis); 3 — мегаладапис Эдвардса (Megaladapis edwardsi).
Мегаладапис Эдвардса (Megaladapis edwardsi Grandidier, 1899) — вид, известный из голоцена южной и юго-западной частей Мадагаскара (местонахождения Амбулисатра (типовое), Ампанихи, Ампоза-Анказоабо, Анавоха, Мономбо, Андрановато, Анкомака, Беавоха, Бетиоки-Толиара, Итамполобе, Ламбохарана, Митохо, Таоламбиби, Тсиандроина, пещера Андрахомана). Синонимы: Megaladapis insignis Forsyth-Major, 1900; Megaladapis brachycephalus Lorenz von Liburnau, 1900; Megaladapis dubius Lorenz von Liburnau, 1900; Palaeolemur destructus Lorenz von Liburnau, 1900.
Самый крупный и специализированный представитель рода. Длина его черепа составляет 27,7-31,7 см, общая длина — 1,3-1,5 м, предполагаемая масса — 70-85 кг. Для этого вида характерны самые крупные в роду коренные зубы (высота M1 — около 18,8 мм), а также наиболее высокие показатели интермембрального (119-120) и самые низкие гумерофеморального (107-110), брахиального (87-90) и крурального (73) индексов. Диафизы плечевой и лучевой костей относительно прямые. Подвздошная кость длинная, расширенная сверху. Коленный сустав сильно вывернут наружу. Бугор на V-й плюсневой кости относительно небольшой.



Смонтированный скелет мегаладаписа Эдвардса (Megaladapis edwardsi) (образец AMNH 15868, слепок). Американский музей естествознания, Нью-Йорк.
Остатки Megaladapis edwardsi в местонахождениях довольно обычны. Некоторые радиоуглеродные датировки позволяют допустить, что он вымер позже остальных видов все еще существовал во время прибытия европейцев в 1504 г.; рассказы местного населения как будто свидетельствуют о том, что он продержался в отдаленных лесных районах вплоть до XVII в.
Мадагаскарский мегаладапис (Megaladapis madagascariensis Forsyth-Major, 1894 (тип) — типовой вид, известный из голоцена южной и юго-западной частей Мадагаскара (местонахождения Амбарарата-Махабо, Амбдисатра, Амбулисатра (типовое), Ампоза-Анказоабо, Анавоха, Мономбо, Анкилитело, Беавоха, Бело-сур-мер, Бемафандри, Итамполобе, Таоламбиби, Тсиандроина, Тсираве, Тсивонохи, пещера Андрахомана). Синоним: Megaladapis filholi Grandidier, 1899.
Размеры наименьшие в роду: череп примерно на 20 % короче, чем у типового вида — 23,5-25 см в длину, высота M1 составляет порядка 14 мм, вес оценивается в 40-45 кг. Индексы пропорций конечностей: интермембральный — 114-118, гумерофеморальный — 112, брахиальный — 94, круральный — 83-90. Плечевая кость в дистальной части расширяется, образуя большой выступ для прикрепления двуглавой мышцы плеча. Диафиз лучевой кости сильно искривлен, также сильно изогнута в продольном плане головка плечевой кости. Крупная пяточная кость с выступающим в срединной плоскости бугром; выдающийся бугор имеется также на V-й плюсневой кости. Суставная головка таранной кости менее уплощена, чем у других видов, и несет более выраженные борозды. Длинный и мощный большой палец стопы далеко отстоит от остальных.

Череп с нижней челюстью мадагаскарского мегаладаписа (Megaladapis madagascariensis).
Мегаладапис Грандидье (Megaladapis grandidieri Standing, 1903) — вид, известный из голоцена северо-центральной части Мадагаскара (местонахождения Ампасамбазимба (типовое), Антсирабе, "Итаси", Морарано-Бетафо). Синоним: Megaladapis gallienii Standing, 1908.
Более крупный, чем Megaladapis madagascariensis, но близкий с ним вид; возможно, оба таксона представляют собой географические разновидности одной клады или морфы. Череп по параметрам напоминает Megaladapis edwardsi, однако коренные зубы находятся в размерном диапазоне Megaladapis madagascariensis (длина черепа составляет 27-30 см; высота М1 — около 15,4 мм). Вес оценивается в 65-75 кг. Щечные зубы относительно небольшие, разделенные широкими диастемами. Индексы пропорций конечностей: интермембральный — 115-117, гумерофеморальный — 113-115, брахиальный — 88-90, круральный — 84-85. Бедренная кость наиболее короткая относительно размеров тела среди всех мегаладаписов, но размеры большой и малой берцовых костей сопоставимы с таковыми у Megaladapis edwardsi (на определенном этапе эти кости Megaladapis madagascariensis даже приписывались еще более крупному лемуру — Archaeoindris fontoynonti). Кривизна проксимальных фаланг по сравнению с другими видами невелика.

Череп с нижней челюстью мегаладаписа Грандидье (Megaladapis grandidieri).
Часть находок определена лишь до рода и обозначена Megaladapis spp. Эти остатки обнаружены на северной оконечности Мадагаскара (местонахождения Ампарихингидро, Анджохибе, Антсируандуха, Андрафиабе). Они обладают морфологическими чертами, общими как с Megaladapis madagascariesis, так и с Megaladapis grandidieri, но их окончательная классификация ввиду фрагментарности затруднена.
Возраст и распространение
Все найденные остатки мегаладаписов — позднеплейстоценовые и голоценовые, происходят из болотных и пещерных отложений Мадагаскара. Обнаружены как отдельные фрагменты костей, так и полные скелеты. Радиоуглеродные датировки, полученные в ходе анализа некоторых остатков мегаладаписа, составили 26150 ± 400 лет (местонахождение Антсируандуха (Анкарана, горный массив на севере острова) и 12760 ± 70 лет (местонахождение Андрафиабе (Анкарана)). Возраст самых молодых остатков из пещеры Анкилителу на северо-западе Мадагаскара методом радиоуглеродного датирования оценивается в 585 ± 65 лет до настоящего времени.

Реконструкция внешнего облика мегаладаписа Грандидье (Megaladapis grandidieri). Автор — Г. Угуэто.
Судя по находкам, Megaladapis grandidieri водился в центральной холмистой части Мадагаскара. Кости Megaladapis edwardsi и Megaladapis madagascariensis часто встречаются в одних и тех же местонахождениях на юге острова; таким образом, эти два вида, занимая одну и ту же экологическую нишу, довольно успешно делили ее между собой. Вместе с тем существуют местонахождения (в том числе Андраумана на крайнем юго-востоке Мадагаскара), где остатки более мелкого Megaladapis madagascariensis почти полностью отсутствуют — возможно, вследствие вытеснения более крупным сородичем.
Образ жизни
Мегаладаписы являлись медлительными животными, проводившими жизнь на деревьях. С помощью хватательных передних и задних конечностей они неторопливо перелезали по стволам и толстым ветвям, способным выдержать их немалый вес, а также совершали короткие вертикальные рывки всем телом. Совершать прыжки с ветки на ветку им не позволяли солидные габариты. При необходимости перейти с одного дерева на другое, мегаладаписы неуклюже передвигались по земле на всех четырех конечностях, возможно, посредством коротких скачков. В прошлом некоторые авторы считали этих лемуров полудревесными животными и указывают на то, что при изобилии подходящего корма и при отсутствии крупных хищников они могли большую часть времени проводить на земной поверхности. Строение полукружных каналов внутреннего уха подтверждает предположение о малоподвижности мегаладаписов. Полагают, что Megaladapis madagascariensis и Megaladapis grandidieri были лучше приспособлены к жизни на деревьях, чем Megaladapis edwardsi, но и он, по-видимому, вел преимущественно древесный образ жизни.

Мегаладапис Эдвардса (Megaladapis edwardsi) на древесных ветвях.
Мегаладаписы были растительноядными, питаясь в основном листьями, которые доставали с помощью удлиненной головы на вытянутой шее. Собирать пищу с большего пространства и подносить ее ко рту помогали более длинные передние конечности. Строение носовых костей позволяют предположить наличие подвижной верхней губы, а отсутствие верхних резцов — длинного языка, служивших для захватывания листвы, что облегчала зубная гребенка в нижней челюсти. О преимущественно лиственном рационе свидетельствуют массивные коренные зубы с высоким рельефом и длинными режущими гребнями, а также характер их микроизноса (царапины и щербины на эмали). На подобный способ питания указывают и маленькие размеры подглазничного отверстия для пучка нервов, ведущих к осязательным рецепторам верхней губы и вибриссам: это отверстие, как правило, больше у животных, в чьем рационе значительную часть составляют фрукты, и меньше у листоядных.
Объемистый желудочно-кишечный тракт были приспособлен к перевариванию значительного количества малопитательной растительности. Секвенирование ядерного генома Megaladapis edwardsi указывает на его конвергентное сходство с листоядными тонкотелыми обезьянами, в генах которых кодирующие белковые продукты участвуют в биоразложении токсинов растений и всасывании питательных веществ. Доля сочных плодов и твердых семян в рационе мегаладаписов была незначительной, в отличие от большинства палеопропитековых (Palaeopropithecidae).
Интересно, что в 1930-е гг. в научной среде существовало предположение, будто мегаладаписы являлись водными животными. Некоторые исследователи считали, что они плавали посредством совершения гребных движений длинными и сильными передними конечностями и были адаптирован к питанию донными моллюсками и ракообразными. При этом сильно вытянутая и уплощенная в передне-заднем направлении подвздошная кость якобы указывает на то, что мегаладаписы были приспособлены к подводным погружениям. В дальнейшем данная гипотеза была отвергнута как не соответствующая новым данным.

Мегаладапис Эдвардса (Megaladapis edwardsi) на поверхности земли. Автор — Р. Учитель.
Значительную роль в жизни мегаладаписов могло играть обоняние: самые мягкие, сочные и молодые листья источают более приятный аромат, чем давно висевшие и набравшие волокнистость. Относительно узкий канал зрительного нерва указывает на сравнительно небольшую резкость зрения и приспособленность к низкому уровню освещения. Головной мозг относительно крупного тела небольшой, объемом около 250 куб. см, что относительно немного для столь крупных приматов.
Естественными врагами мегаладаписов были лазающие хищники — в первую очередь, вымершая гигантская фосса (Cryptoprocta spelea), жертвами которой становились преимущественно молодые особи. Кроме того, лемуры, спускавшиеся на землю, чтобы перейти от дерева к дереву, могли становиться добычей наземных хищников. В частности, некоторые найденные кости мегаладаписов несут следы зубов крокодилов.
Установлено, что постоянные зубы у мегаладаписов прорезались на очень ранней стадии индивидуального развития (хотя и не настолько ранней, как у палеопропитеков (Palaeopropithecus)). Раннее развитие постоянных зубов и небольшой объем мозга у субфоссильных лемуров (включая мегаладаписов) рассматриваются как признаки того, что эти приматы рано переходили к самостоятельной, независимой от родителей жизни и, возможно, рано достигали половой зрелости. При этом исследования генома показывают низкий уровень генетического разнообразия мегаладаписов, что свидетельствует о небольших размерах их популяций.
Вымирание
По вопросу о причине исчезновения мегаладаписов велись долгие споры. Один из возможных факторов, негативно отобразившийся на их численности — длительный засушливый период, приведший к сокращению площади тропических лесов, служивших листоядным лемурам местообитаниями (считается, что пик засухи пришелся на XIV в.). Однако представляется куда более вероятным, что к исчезновению мегаладаписов и почти трети прочих видов мадагаскарских лемуров в первую очередь привели преднамеренные пожары для сведения лесов под сельскохозяйственные нужды и непосредственное истребление малайскими иммигрантами, впервые прибывшими на остров приблизительно в начале нашей эры (впрочем, в разных источниках эта дата колеблется от 500 г. до н. э. до 500 г. н. э.). В любом случае, кости мегаладаписов иногда находят в культурных слоях за сотни километров от границы леса и на них порой присутствуют следы кухонной обработки. Медлительным гигантам могли угрожать и размножившиеся одичавшие свиньи.

Скульптурная реконструкция мегаладаписа Эдвардса (Megaladapis edwardsi) на поверхности земли. Автор — К. Флеров.
Сегодня большинство авторов сходится на том, что мегаладаписы окончательно исчезли в XIII-XV вв. Вероятнее всего, европейские поселенцы их уже не застали. Впрочем, они вполне могли услышать рассказы об этих необычных лемурах из уст местных жителей. Так, исследователь Э. де Флакур в «Истории Великого острова Мадагаскар» (1661 г.) писал: "Третретретре — это животное размером с двухгодовалого теленка. У него круглая голова, а лицо похоже на человеческое. Однако передние и задние конечности сходны с обезьяньими. У него курчавый мех, короткий хвост и почти человеческие уши. Это животное ведет уединенный образ жизни. Местное население очень боится его, и поэтому при встрече и та и другая сторона спасаются бегством". Впрочем, некоторые авторы относят вышеприведенную цитату не к мегаладаписам, а к другим крупными вымершим лемурам — палеопропитекам (Palaeopropithecus). Корни образа мегаладаписов они ищут в другом животном из рассказов мальгашей — токандии, "крупном, живущем на деревьях звере, способном совершать прыжки и имеющем лицо, лишь отдаленно схожее с человеческим" (ведь морда мегаладаписов была сильно вытянута, в отличие от палеопропитеков).
Ссылки
Мегаладаписы (Википедия) перейти
Megaladapis (Paleobiology Database) перейти
Литература
Дробышевский С. Достающее звено. В 2 т. Т. 1. Обезьяны и все-все-все. — Corpus, 2017
Каландадзе Н. Н., Шаповалов А. В. Судьба мегафауны наземных экосистем Мадагаскарской зоогеографической области в позднем антропогене.
Ламберт Д. Доисторический человек: Кембриджский путеводитель. — Л.: Недра, 1991
Мадагаскар / Науч. ред. Б. Д. Васильев. — М.: Прогресс, 1990
Alcover J. A., Campillo X., Macias M., Sans A. Mammal Species of the World: Additional Data on Insular Mammals. American Museum Novitates 3248, 1998
Cachel S. Fossil Primates. — Cambridge, UK: Cambridge University Press, 2015
Cenozoic mammals of Africa / Ed. by Werdelin L., Sanders W. J. — University of California Press, 2010
Ciochon R. L, Fleagle J. G. Primate evolution and human origins. — Hawthorne, NY: Aldine de Gruyter, 1987
Eberhart G. M. Mysterious Creatures: A Guide to Cryptozoology. — ABC-CLIO, 2002
Encyclopedia of Human Evolution and Prehistory / Ed. by Delson Е., Tattersall I., Van Couvering J., Brooks A. S. — Taylor & Francis, 2000
Fleagle J. G. Primate Adaptation and Evolution (2rd ed.). — Academic Press, 1998
Forsyth-Major C. I. On Megaladapis madagascariensis, an extinct gigantic lemuroid from Madagascar; with remarks on the associated fauna, and on its geological age. Philosophical Transactions of the Royal Society B 185, 1894
Forsyth-Major C.I. Extinct Mammalia from Madagascar. I. Megaladapis insignis, sp.n. Philosophical Transactions of the Royal Society 193, 1900
Godfrey L. R. The International Encyclopedia of Primatology / Ed. by Fuentes A. — Wiley Blackwell, 2017
Goodman M. S., Jungers W. L. Extinct Madagascar: Picturing the Island's Past. — The University of Chicago Press, 2014
Grzimek's animal life encyclopedia. Volumes 12-16 (2rd ed.) / Ed. by Hutchins M., Kleiman D. G., Geist V., McDade M. C. — Farmington Hills, MI: Gale Group, 2004
Herrera J. P., Dávalos L. M. Phylogeny and Divergence Times of Lemurs Inferred with Recent and Ancient Fossils in the Tree. Systematic Biology 65 (5), 2016
Karanth K. P., Delefosse T., Rakotosamimanana B., Parsons T. J., Yoder A.D. Ancient DNA from giant extinct lemurs confirms single origin of Malagasy primates. Proceedings of the National Academy of Sciences. 102 (14), 2005
Lemurs: Ecology and Adaptation / Ed. by Gould L., Sauther M. L. — Springer Science & Business Media, 2006
Mittermeier R. A., Louis E. E., Richardson M., Schwitzer C., Langrand O., Rylands A. B., Hawkins F., Rajaobelina S., Ratsimbazafy J., Rasoloarison R., Roos C., Kappeler P. M., MacKinnon J. Lemurs of Madagascar (3rd ed.). — Conservation International, 2010
New Directions in Lemur Studies / Ed. by Rakotosamimanana B., Rasamimanana H., Ganzhorn J., Goodman S. M. — Springer Science+Business Media, 1999
Nowak R. M. Walker's Primates of the World. — Baltimore, MD: The Johns Hopkins University Press, 1999
Perez V.R., Godfrey L.R., Nowak-Kemp M., Burney D.A., Ratsimbazafy J., Vasey N. Evidence of early butchery of giant lemurs in Madagascar. Journal of Human Evolution 49 (6), 2005
Reconstructing Behavior in the Primate Fossil Record / Ed. by Plavcan J. M., Kay R. F., Jungers W. L., van Schaik C. P. — Springer Science & Business Media, 2012
Szalay F., Delson E. Evolutionary history of the Primates. — New York: Academic Press, 1979
Tattersall I. The Primates of Madagascar. — New York: Columbia University Press, 1982
The Natural History of Madagascar / Ed. by Goodman S. M., Benstead J. P. — University of Chicago Press, 2003
The Primate Fossil Record / Ed. by Hartwig W. C. — Cambridge: Cambridge University Press, 2002
Wunderlich R. E, Simons E. L., Jungers W. L. New pedal remains of Megaladapis and their functional significance. American Journal of Physical Anthropology 100 (1), 1996