Мегантропы — Meganthropus
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Суперотряд Эвархонтоглиресы (Euarchontoglires)
Грандотряд Эвархонты (Euarchonta)
Миротряд Приматообразные (Primatomorpha)
Отряд Приматы (Primates)
Подотряд Сухоносые приматы (Haplorrhini)
Инфраотряд Настоящие обезьянообразные (Simiiformes)
Парвотряд Узконосые обезьяны (Catarrhini)
Клада Настоящие узконосые обезьяны (Eucatarrhini)
Надсемейство Гоминоиды (Hominoidea)
Семейство Человекообразные (Hominidae)
Подсемейство Понгины (Ponginae)
Род †Мегантропы (Meganthropus)
Вид †Meganthropus paleojavanicus
Мегантропы (Meganthropus Koenigswald, 1950) — монотипический род подсемейства понгин (Ponginae Elliot, 1913), происходящий из раннего плейстоцена Азии.
Реконструкция внешнего облика мегантропа (Meganthropus paleojavanicus). Автор — Дж. Кнюппе.
Синонимы: Homo Linnaeus, 1758 (частью); Pongo Lacépède, 1799 (частью); Pithecanthropus Dubois, 1894 (частью); Australopithecus Dart, 1925 (частью).
Единственный установленный вид — Meganthropus paleojavanicus Koenigswald, 1950 (тип). Его остатки обнаружены на острове Ява, Индонезия. Они датируются ранним плейстоценом. Абсолютный возраст в настоящее время оценивается приблизительно в 1,15-0,7 млн. лет, более точная датировка затруднена из-за условий захоронения и обнаружения остатков. Синонимы: Pithecanthropus dubius; Australopithecus palaeojavanicus; Homo palaeojavanicus; Homo erectus palaeojavanicus; Pongo sp.
История изучения
Первые находки (нижнечелюстные фрагменты Сангиран 5 и 6) были совершены в 1939 и 1941 гг. в местонахождении Сангиран в долине реки Соло в центральной части острова Ява голландским геологом Г.Г.Р. фон Кенигсвальдом. Обнаруженные остатки привлекали внимание своей толщиной и массивностью, поэтому автор открытия предложил для них название Meganthropus palaeojavanicus. Родовое наименование образовано древнегреческими словами μέγας — "большой" и ἄνθρωπος — "человек", а видовой эпитет в переводе с латыни означает "древнеяванский". Такое название название указывает на принадлежность найденных образцов крупной форме древнего человека и географическое место обнаружения. Впервые оно было употреблено автором открытия в письме за 1941 г. и впоследствии использовалось рядом исследователей, хотя первое формальное описание таксона под этим названием было опубликовано Кенигсвальдом лишь в 1950 г.
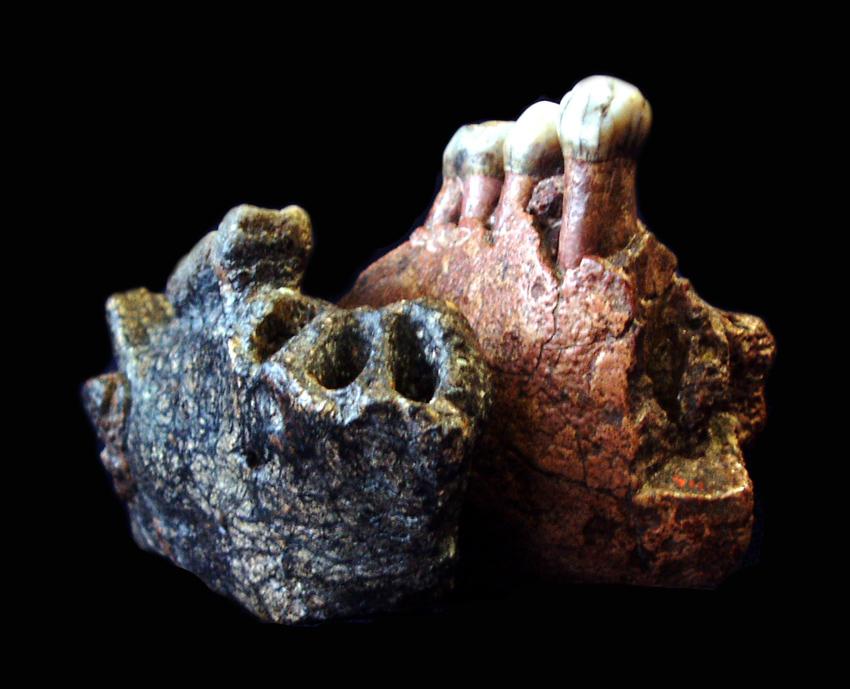
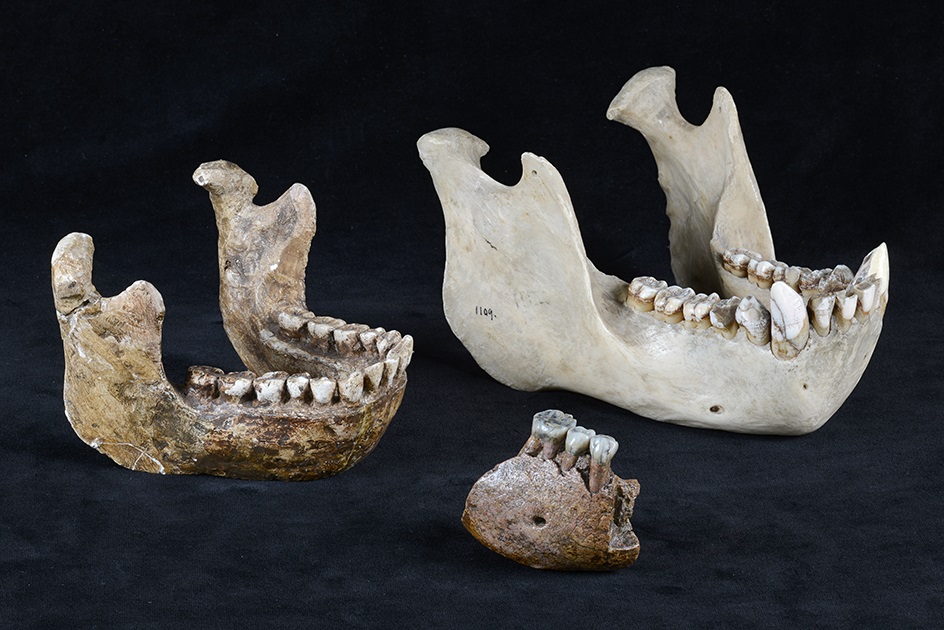
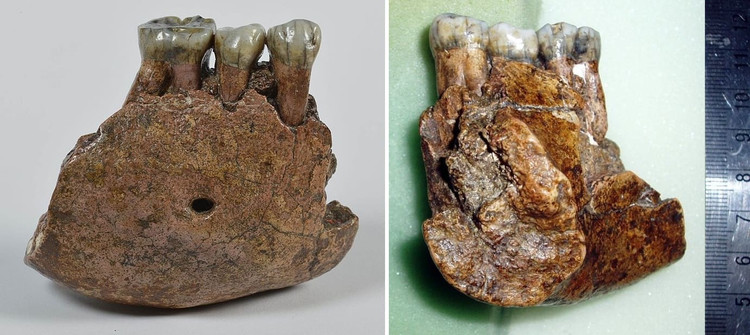
Нижнечелюстные фрагменты мегантропа (Meganthropus paleojavanicus) Сангиран 5 и 6.
Характерная черта мегантропа — значительная массивность имеющегося материала, что может свидетельствовать о сравнительно крупных размерах тела. Немецкий исследователь Ф. Вейденрейх построил на этих находках оригинальную гипотезу о гигантизме предков человека и местной линии эволюции гоминид, обнародовав первые результаты своих изысканий уже в 1942 г. Он полагал, что промежуточный по размерам мегантроп (которого он именовал яванским Голиафом) был потомком гигантопитека и стал предком для питекантропа, а через него — представителей желтой расы современного человека. Кенигсвальд присоединялся к этому взгляду без особой уверенности. Немецкий ученый Г. Вейнерт в 1951 г. попытался оживить гигантоидную теорию. Впоследствии данное заключение, полифилетическое по своей сути, было целиком отвергнуто наукой. Согласно другой точке зрения, островная изоляция на протяжении сотен тысяч лет способствовала увеличению общих размеров мегантропа.
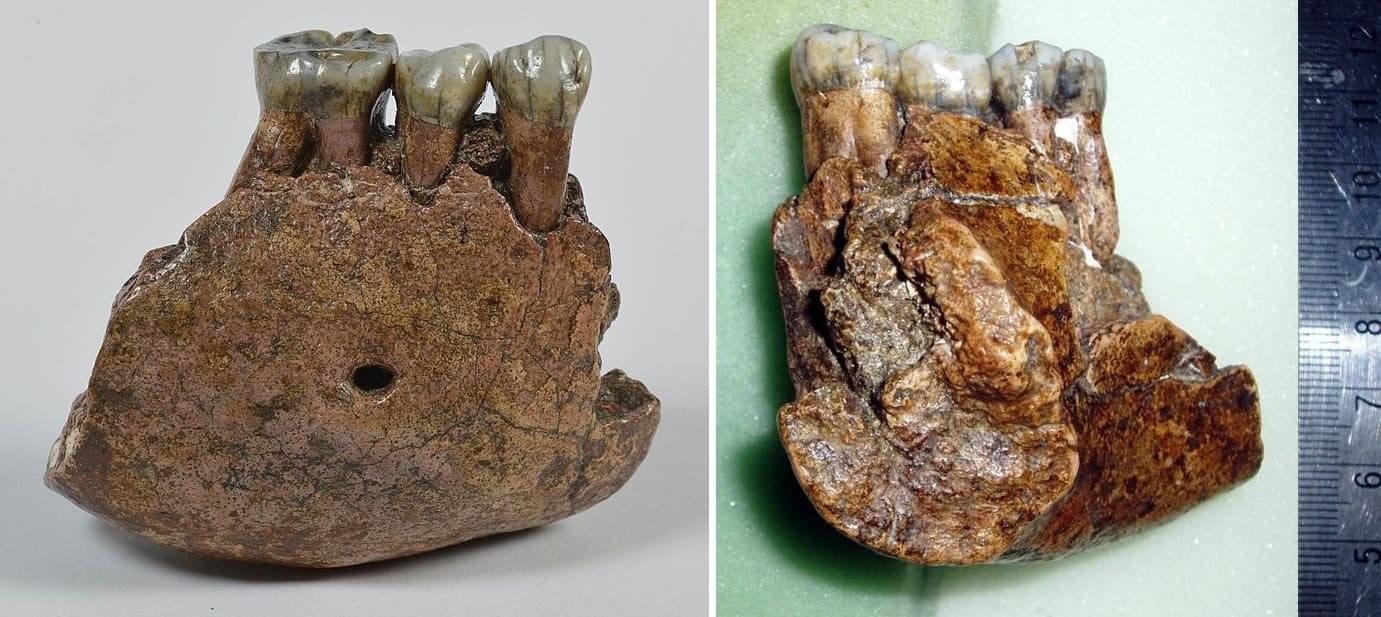
Нижнечелюстной фрагмент мегантропа (Meganthropus paleojavanicus) Сангиран 6.
В 1939 г. Коль-Ларсен открыл в районе озера Виктория на востоке Африки среди прочих ископаемых остатков кусок правой верхней челюсти с 2 предкоренными зубами, а в нескольких километрах от места этой находки обнаружил верхний коренной такой же окраски, сохранности и подходящего размера. Эти остатки Вейнерт в 1950 г. включил в род мегантропов, но в качестве другого вида — мегантропа африканского (Meganthropus africanus). После открытия черепа австралопитека в Сварктрансе, ЮАР, в 1948 г. (образец SK48) он некоторое время также обозначался в научной литературе как Meganthropus africanus. Однако вскоре выяснилась принадлежность африканских находок к парантропу массивному (Paranthropus robustus), а предыдущее название было признано его младшим синонимом.
В 1953 г. южноафриканский палеоантрополог Дж. Т. Робинсон рассматривал Meganthropus palaeojavanicus (на основе фрагмента нижней челюсти Сангиран 6) в качестве азиатского представителя австралопитеков. Эта позиция получила развитие в работах некоторых специалистов, например Кенигсвальда (1973 г.), Крантца (1975 г.), Орбан-Сегебарта и Прокуреура (1983 г.). Одно время даже велась дискуссия о том, к какой форме австралопитека мегантроп ближе — изящной или массивной. Однако в целом сторонников у данной гипотезы нашлось немного и ее убедительных подтверждений на текущий момент не существует.
Наличие подбородочной ости (костного выступа, служащего для прикрепления мышц языка) свидетельствует о том, что мегантроп мог владеть членораздельной речью. Впрочем, надежных сопутствующих следов культуры (изготовления орудий труда, разведения огня и т. п.) для него не обнаружено. Поэтому было высказано предположение, что мегантропы жили небольшими семейными группами, кочуя в поисках подходящей пищи растительного и животного происхождения.
В 1950-е гг. российский антрополог М.А. Гремяцкий осуществил монтаж нижней челюсти мегантропа с верхней челюстью Сангиран 4, принадлежавшей моджокертскому подвиду выпрямленного человека (Homo erectus modjokertensis), показав при этом, что обе челюсти хорошо гармонируют друг с другом. Кроме того, места находок и датировки мегантропа те же самые, что и у Homo erectus modjokertensis. Помимо прочего, последний отличался значительной массивностью скелета, но имел при этом вполне средний рост. Как следствие, Гремяцкий сделал вывод, что мегантроп вряд ли заслуживает выделения в отдельный вид, а тем более род.
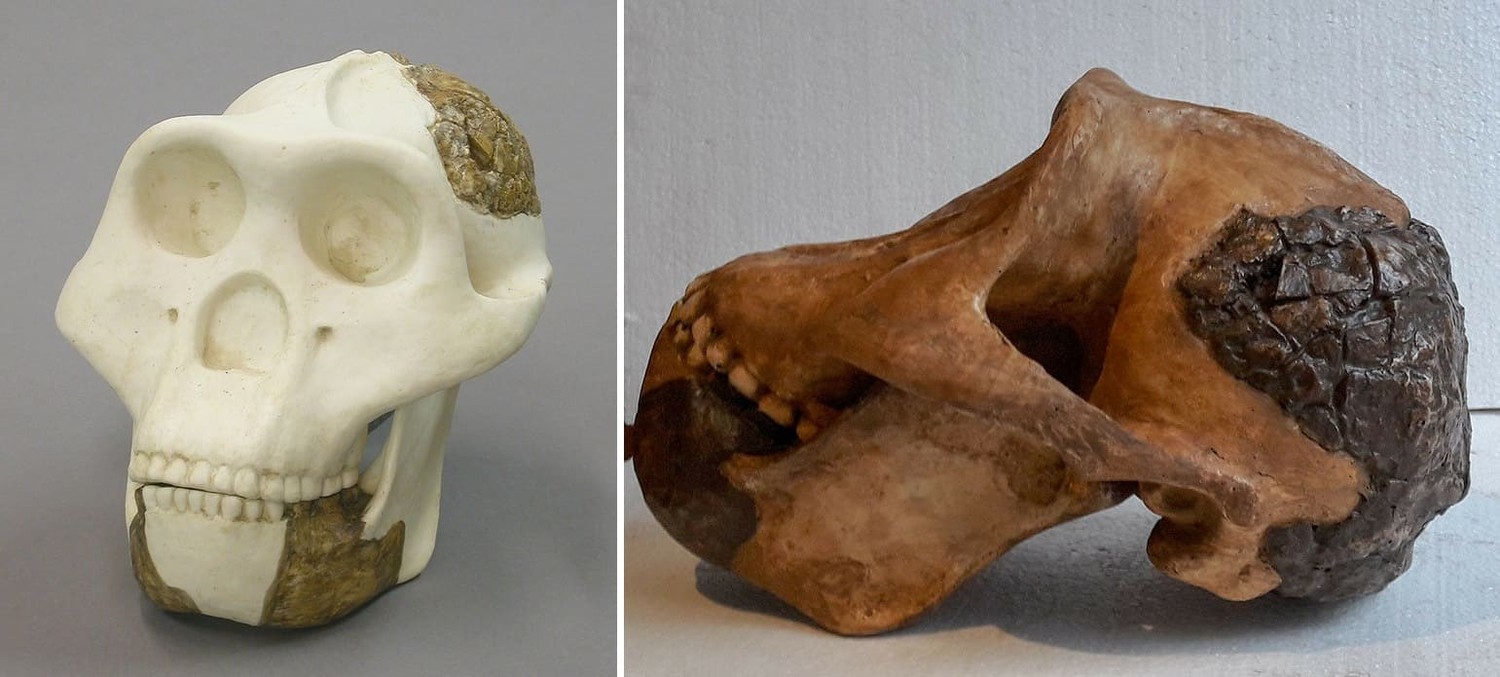
Человекоподобная реконструкция черепа мегантропа (Meganthropus paleojavanicus), основанная на нижнечелюстном фрагменте Сангиран 6 и задней части черепа Сангиран 31. Автор — Г. Крантц.
В дальнейшем сама возможность существование отдельного таксона Meganthropus paleojavanicus неоднократно оспаривалась различными исследователями. Большинство специалистов (М.А. Гремяцкий, У. Ле Гро Кларк, Ж. Пивето и др.) сошлись во мнении, что никакие морфологические признаки не позволяют говорить о выделении мегантропа из круга питекантропов (ныне — Homo еrectus). Некоторые авторы полагали, что особенно массивные остатки, традиционно приписываемые мегантропу, просто принадлежат самцам Homo еrectus. При таком их истолковании многое зависит от того, какой именно диапазон внутривидовой изменчивости принимать в качестве нормы для "обычного" Homo еrectus. И хотя род Meganthropus фактически был упразднен, это название осталось неформальным обозначением для ряда яванских находок.
Однако, с этим согласилась не вся научная общественность. Например, М.Ф. Нестурх в 1954 г. причислил мегантропа к человекообразным обезьянам. С. Сартоно, хотя и считал мегантропа близким к Homo еrectus, выделил его в отдельный вид человек древнеяванский (Homo paleojavanincus). Иногда признавалось существование самостоятельного подвида Homo еrectus palaeojavanicus. Крамер и Кенигсберг (1994 г.), Вольпофф (1999 г.), Картмилл и Смит (2009 г.) оспаривают эту точку зрения, приводя доводы в пользу сильного сходства между более ранними и поздними яванскими образцами и отсутствия различий между видами или подвидами.
В 2009 г. Р. Циочон предложил возможность существования в плейстоцене Явы не установленного пока примата, хотя все еще считал мегантропа близким к Homo еrectus. В 2019 г. немецкий ученый К. Занолли и его коллеги использовали анализ окклюзионных отпечатков зубов для реконструкции жевательной кинематики и различные морфометрические подходы, основанные на микротомографии, чтобы исследовать внутренние зубные структуры мегантропа. Оказалось, что его зубы существенно отличаются как от таковых человека прямоходящего, но демонстрируют значительное подобие зубам вымерших понгин люфенгпитеков (Lufengpithecus) и сивапитеков (Sivapithecus) в отношении распределения толщины эмали, поверхности и положения зубчатых выступов во внутренней части зубных коронок, а также сходный с орангутанами (Pongo) характер износа жевательных поверхностей. Таким образом, концепция "загадочной обезьяны" подтвердилась: род и вид Meganthropus paleojavanicus были возвращены в научный оборот, но не как часть эволюционной линии рода Homo.
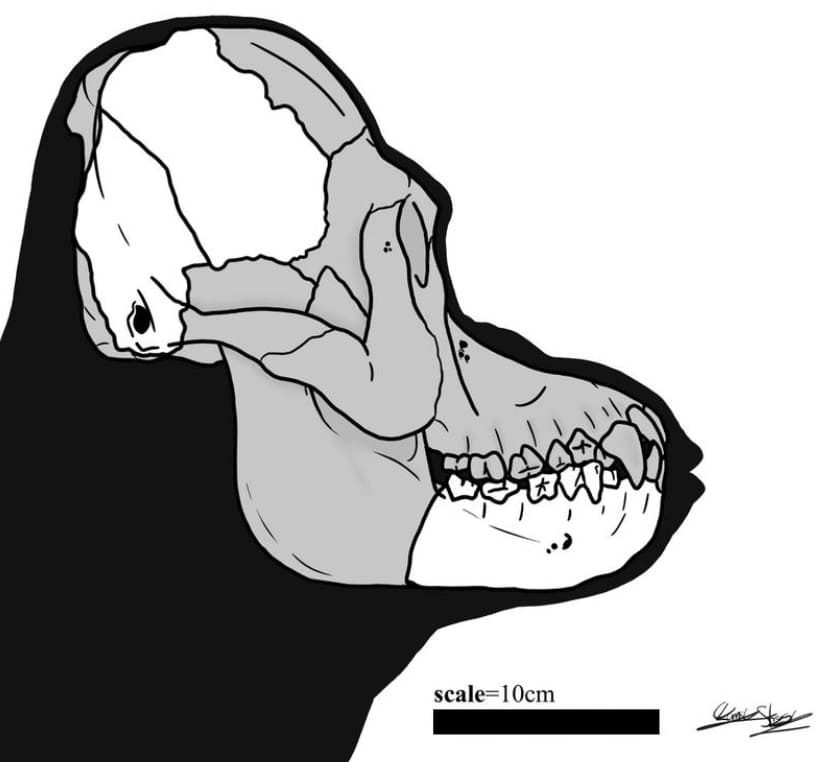
Прорисовка реконструированного черепа с нижней челюстью мегантропа (Meganthropus paleojavanicus).
Находки
Обнаружено сравнительно немного окаменелых остатков, принадлежащих Meganthropus paleojavanicus. К сожалению, все они отличаются фрагментарностью. В большинстве своем это изолированные зубы и части нижних челюстей. Вероятно, имеются также части лицевого и мозгового черепа.
Триниль 11620 и Триниль 11621. 2 крупных коренных зуба, найденные Э. Дюбуа еще в 1891 г. Автор открытия отнес их к
Pithecanthropus erectus, однако последние исследования опровергают их гомининное происхождение. Судя по всему, эти зубы принадлежали мегантропу.
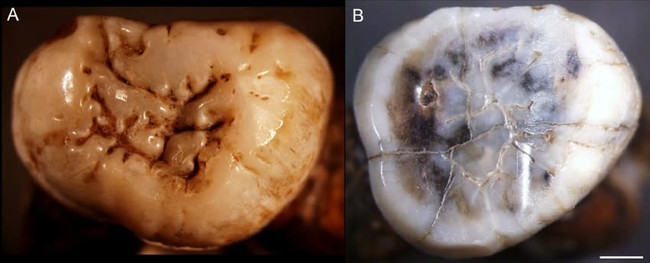
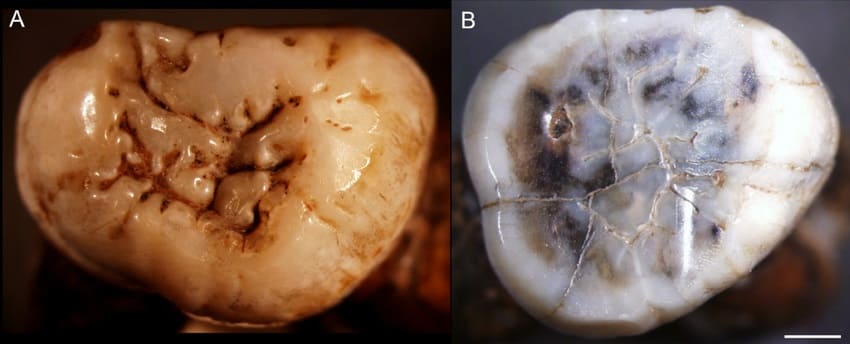
Коренные зубы мегантропа (
Meganthropus paleojavanicus) Триниль 11620 (А) и Триниль 11621 (В). Длина масштабной линейки — 2 мм.
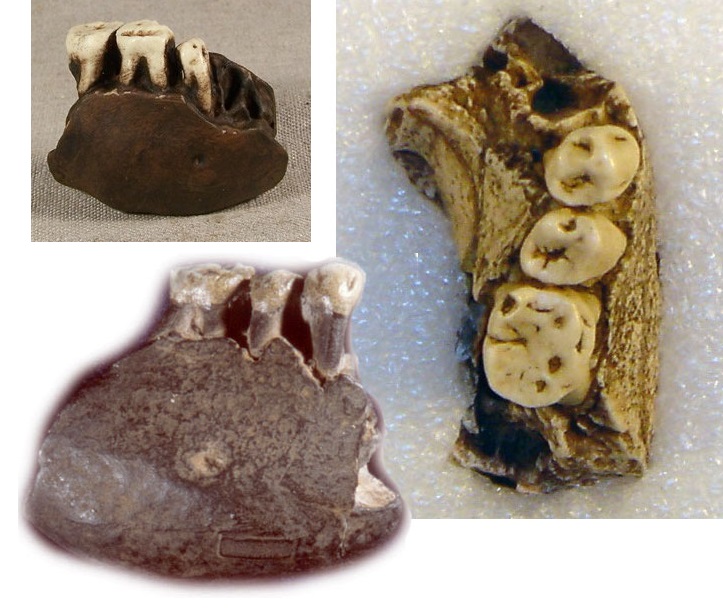
Питекантроп С / Сангиран 5. Обломок правой половины тела нижней челюсти, найденный весной 1939 г. в раннеплейстоценовых слоях формации Пуканган, содержащих остатки фауны Джетис. Оценочный возраст вмещавших образцы отложений составил 1,67 млн. лет. Найденный фрагмент охватывает область от промежутка между m2 и m3 до уровня i2. В челюсти сохранились сильно стертые p2, m1 и m2. Образец очень велик и по величине тела челюстной кости приближается к таковой у орангутана. Толщина кости в области симфиза равна 19 мм, превышая толщину кости у шимпанзе (максимум 17,5 мм) и уступая горилле (в среднем 23 мм). Не изучая специально морфологии этого фрагмента, Кенигсвальд был склонен отнести его к остаткам питекантропов. Так как основание тела челюсти узкое, как у антропоидов и не имеет ямки двубрюшной мышцы, Вейденрейх склонялся к мысли, что нижненечелюстной фрагмент принадлежит человекообразной обезьяне (возможно, орангутану). Из-за крупных размеров эти остатки в настоящее время обычно относят к мегантропу.

Нижнечелюстной фрагмент мегантропа (Meganthropus paleojavanicus) Сангиран 5.
Мегантроп A / Сангиран 6. Нижнечелюстной фрагмент, обнаруженный местным сборщиком окаменелостей в конце апреля 1941 г. и переданный Кенигсвальду. Считается, что данный образец происходит из тех же слоев Пуканган с фауной Джетис. Незадолго перед этим них уже была извлечена верхняя челюсть Питекантроп IV / Сангиран 4, определяемая как Homo erectus modjokertensis. Сохранилась часть правой стороны тела нижней челюсти с альвеолами i1, i2 и c, а также целыми р1, р2 и m1. Эта челюсть гораздо больше предыдущей и уступает по размерам только нижним челюстям очень крупных самцов горилл. Наибольшая высота кости в области симфиза составляет 48 мм, толщина — 25,5 мм (у самца гориллы — до 31 мм).
Позднее было показано, что, судя по альвеолам, резцы, вероятно, были развиты относительно слабо, контрастируя с мощными щечными зубами, т. е. имело место такое же соотношение величины передних и задних зубов, как у австралопитеков. Исходя из этого, образец был обозначен Дж. Т. Робинсоном как парантроп древнеяванский (Paranthropus paleojavanicus). Оба предкоренных мегантропа имеют по 1 корню; у австралопитеков по большей части — 2 корня, хотя встречается и 1 корень. Наконец, асимметрия бугорков и бороздок, которой (по Робинсону) мегантроп якобы особенно сходен с австралопитеками, очень обычна на коронках нижних предкоренных современного человека.
Нижнечелюстной фрагмент мегантропа (Meganthropus paleojavanicus) Сангиран 6.
Во время Второй мировой войны Кенигсвальд был захвачен японцами и интернирован в концентрационный лагерь, но до этого он успел отправить слепки найденных фрагментов для дальнейшего изучения немецкому анатому Ф. Вейденрейху в Американский музей естественной истории в Нью-Йорке, США. Сам Кенигсвальд полагал, что образцы Сангиран 5 и 6 принадлежат новому крупному виду древних людей и представляют соответственно самку и самца. Детально сравнить морфологию зубов обоих фрагментов не удалось вследствие плохой сохранности первого.
Вейденрейх придерживался иного мнения и описал Сангиран 5 как Pithecanthropus dubius, поскольку он сомневался, что данный фрагмент принадлежал человеку (питекантропы на тот момент частью исследователей собственно людьми не признавались). Тем не менее, он согласился с тем, что Сангиран 6 являлся человеком и принял его название "мегантроп древнеяванский" (Meganthropus paleojavanicus), которое было предложено Кенигсвальдом в сопроводительном письме. Это наименование родилось под впечатлением от непривычно большего размера и массивности фрагмента Сангиран 6. Включавшая его челюсть была приблизительно той же высоты, что и у гориллы, но имела гораздо большую толщину. Зубы по сравнению с челюстью сравнительно невелики. Вейденрейх исключал, что размеры челюсти являются патологическим признаком, также он отверг возможность акромегалии или иного уродства владельца челюсти.
Мегантроп B / Сангиран 8. Передняя часть нижней челюсти с единственным m3 была найдена в 1952 г., а в дальнейшем выкуплена у местных жителей и описана доктором П. Марксом из Бандунгского университета в 1953 г. Данный образец происходит из слоев более молодой формации Бапанг. Его размеры и строение сопоставимы с предыдущим образцом, но он еще до окаменения получил значительные повреждения — был раздавлен зубами крупного крокодила. Японские и индонезийские исследователи пришли к выводу, что образец принадлежал взрослой особи и имел определенные морфологические особенности, отсутствующие у "обычного" Homo erectus. В то же время он обладает чертами, характерными для более ранней находки Мегантроп A / Сангиран 6.
Нижнечелюстной фрагмент мегантропа (Meganthropus paleojavanicus) Сангиран 8.
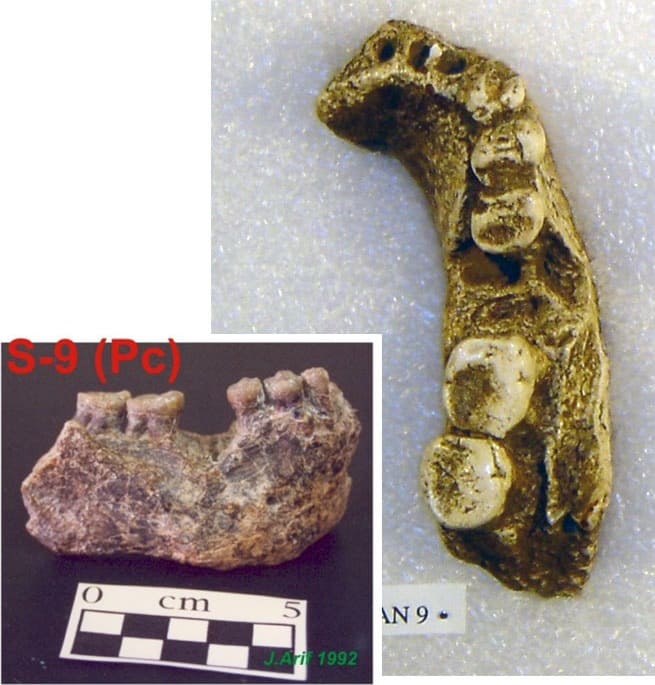
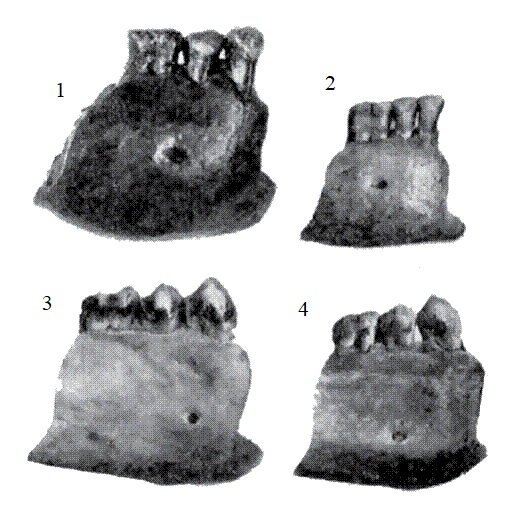
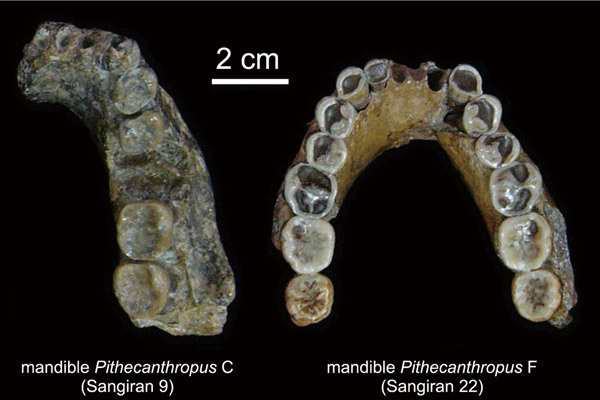
Мегантроп C / Сангиран 9. Эта находка была совершена С. Сартоно в ноябре 1960 г. в Тегопати в нижних слоях формации Пуканган. Она представляет собой массивный фрагмент нижней челюсти с с, р1, р2, m2 и m3.
Нижнечелюстной фрагмент мегантропа (Meganthropus paleojavanicus) Сангиран 9.
Сангиран 33 / BK 7905. Нижнечелюстной фрагмент, обнаруженный в 1979 г. и обладающий некоторыми общими с предыдущей находкой характерными признаками. Однако ее принадлежность мегантропу отвергается рядом исследователей.
Мегантроп D / ? Сангиран 70. Эти фрагмент нижней челюсти и ее ветвь были приобретены С. Сартоно в 1993 г. и датированы возрастом около 1,4-0,9 млн. лет. Часть ветви сильно повреждена, фрагмент же нижней челюсти сохранился сравнительно хорошо, хотя сохранность зубов и оставляет желать лучшего. Размеры челюсти несколько меньше, чем у образца Мегантроп А / Сангиран 6, но по форме она очень похожа на него. По заключению Сартоно, Тайлера и Крантца, представляется весьма вероятным, что Meгантропы A и D принадлежат одной и той же разновидностью гоминида, кем бы он не оказался при дальнейшем уточнении.
Мегантроп I / Сангиран 27. Часть массивного лицевого скелета, найденная местным жителем во время копания оросительной канавы близ деревни Нгампон к югу от Сангирана в марте 1978 г. Описаны Д. Тайлером как принадлежащие Meganthropus paleojavanicus в противоположность Homo erectus. А.Ц. Дурбанд оспорил эти выводы и отнес череп к человеку выпрямленному, показав, что его параметры находится в пределах внутривидовой изменчивости последнего.
Нижняя часть лицевого скелета мегантропа (Meganthropus paleojavanicus) Сангиран 27.
Мегантроп II / Сангиран 31. Задняя часть мозгового черепа, обнаруженная при подобных же обстоятельствах и описанная С. Сартоно в 1982 г. Находка происходит из слоя с остатками фауны Джетис, ее возраст был определен приблизительно в 1,8 млн. лет. Как и большинство других окаменелостей такого рода, она сильно повреждена, но ее относительная полнота в некоторой мере компенсирует невысокий уровень сохранности. Д. Тайлер пришел к выводу, что параметры черепа находятся вне обычного диапазона Homo erectus. Он более низкий и широкий и имеет стенки толще, чем любой восстанавливавшийся ранее череп человека выпрямленного. Особенно примечательным для данного образца является наличие двойного сагиттального гребня — деталь анатомического строения, неизвестная у "обычного" Homo erectus. Затылочный бугор очень массивный. Расчетная емкость мозговой коробки составляет 800-1000 куб. см. С момента обнародования в 1993 г. и по сей день реконструкция Тайлера образца Сангиран 31 принята большинством авторитетных специалистов и считается достаточно правдоподобной.


Задняя часть мозгового черепа мегантропа (Meganthropus paleojavanicus) Сангиран 31.
После изучения этого фрагмента и его последующей реконструкции Э. Крамер пришел к выводу, что данный образец весьма схож с Питекантропом IV / Сангираном 4, определенного как человек прямоходящий моджокертский (Homo erectus modjokertensis). А.Ц. Дурбанд указывает, что хотя по ряду анатомических особенностей Мегантропа II и отличается от "обычного" Homo erectus, в целом он не выходит за пределы его внутривидовой изменчивости. Предположение Г. Крантца о том, что Сангиран 31 является гигантским представителем человека умелого (Homo habilis), весьма спорно.
Мегантроп III. Находка представляет собой фрагмент задней части черепа. Принадлежность этого образца к мегантропу является довольно сомнительной. Описавший его в 1996 г. Д. Тайлер указывал, что затылочный угол всего черепа должен был составлять около 120 °, что, по его мнению, находится вне известного диапазона изменчивости Homo erectus, затылок которого гораздо более изогнут. В то же время морфологический анализ, проведенный А.Ц. Дурбандом, помещает образец в рамках "обычного" Homo erectus. Также существует мнение, что данная находка вообще не является частью черепа гоминида.
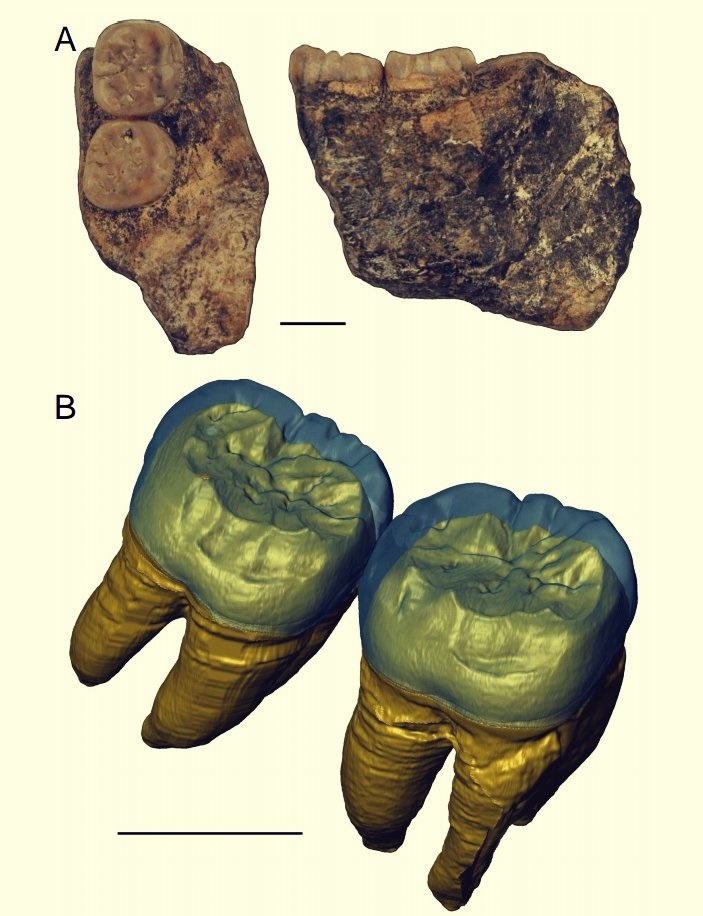
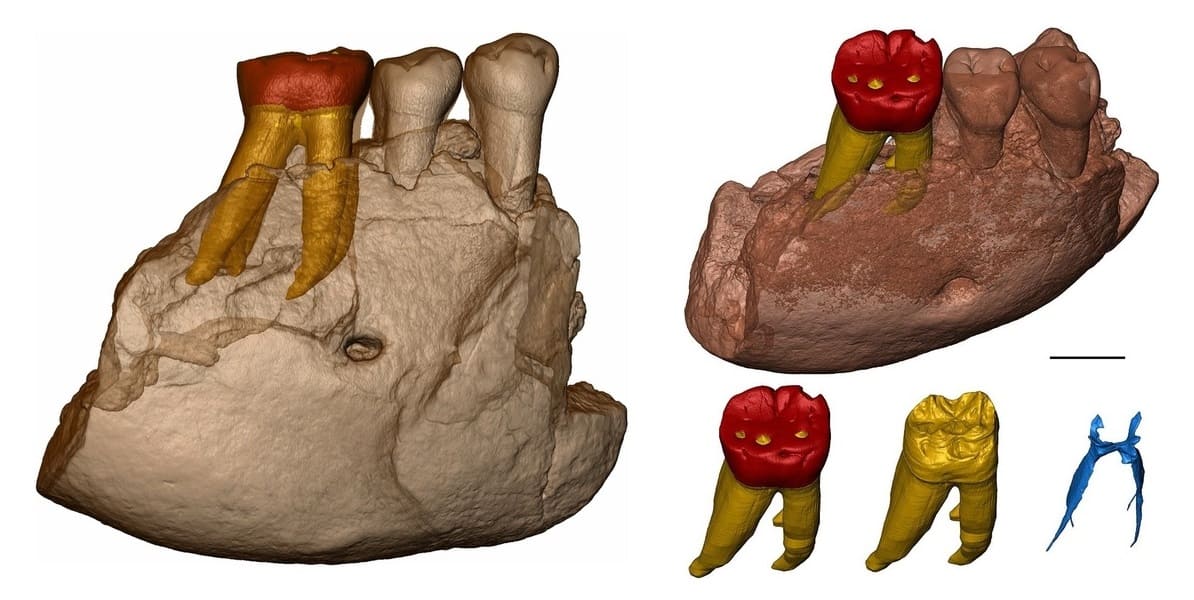
Арджуна 9. Нижнечелюстной фрагмент с 2 коренными зубами, обнаруженный в мае 1988 г. возле Нгебунга, к северу от Сангирана.
Нижнечелюстной фрагмент мегантропа (Meganthropus paleojavanicus) Арджуна 9 и компьютерная модель его коренных зубов.
Помимо вышеперечисленных образцов, имеется ряд разрозненных зубов (FS-77, SMF-8855, SMF-8864, SMF-8879 и др.), приналлежавших мегантропу. Ввиду поверхностного сходства многие из них ранее приписывались орангутану или выпрямленному человеку.
Строение и функции
Нижняя челюсть мегантропа чрезвычайно крупная и массивная, превышающая по этим показателям других гоминид, кроме гигантопитеков, самцов горилл, некоторых австралопитеков и Дманиси 2600. Тогда как у человекообразных обезьян нижняя челюсть имеет наибольшую высоту в симфизе, в данном случае наибольшая высота фиксируется примерно на уровне m1. Если же измерять толщину тела нижней челюсти в области симфиза, то наибольшие размеры у человекообразных обезьян получаются выше поперечного валика; у мегантропа, как у гоминин, максимальная толщина приходится ниже этого валика. В области коренных зубов нижняя челюсть по диаметру даже больше челюсти гориллы.
Подбородочного выступа нет, а симфиз челюсти ровный, слабо скошен назад, закруглен снизу. Передняя сторона симфиза наклонена к горизонтальной плоскости под углом приблизительно 60°. Этот показатель близок к таковому многих австралопитеков и поздних архантропов, больше чем у человекообразных обезьян (45-55°) и меньше чем у ранних архантропов и неандертальцев (70-80°); у современных людей в связи с развитием подбородочного выступа угол прямой или тупой.
Нижнечелюстной фрагмент мегантропа (Meganthropus paleojavanicus) Сангиран 6 (в центре), а также нижние челюсти прямоходящего человека (Homo erectus) (слева) и орангутана (Pongo pygmaeus) (справа).
В отличие от большинства челюстей современного человека, тело челюсти мегантропа не расширяется и не утолщается в районе симфиза. Поверхность щечной стороны довольно гладкая. Косая линия (linea obliqua), обычно дающая 2 ветви (torus lateralis superior, torus marginalis), выражена здесь очень слабо, так что бороздка между этими ветвями (sulcus intertoralis) не прослеживается. Нижний край челюсти представляет довольно широкую округленную поверхность; его форма отличается от формы, свойственной гомининам, у которых обычно наблюдается постепенное расширение этого края от уровня m3 к симфизу.
Подбородочное отверстие (foramen mentale) мегантропа одиночное (у человекообразных обезьян оно двойное-тройное), широкое и открывается сбоку примерно на середине высоты челюсти, как у гоминин (у обезьян оно лежит значительно ниже середины высоты горизонтальной ветви). По своему положению отверстие делит тело челюсти на 2 равные части — базальную и альвеолярную — в отличие от человекообразных обезьян, у которых резко преобладает альвеолярная часть, занимая в среднем 2/3 высоты. Подбородочным отверстием заканчивается нижнечелюстной канал (canalis mandibularis), который, как и у современного человека, на большей части своего протяжения лежит ниже середины высоты горизонтальной ветви.
Нижнечелюстной фрагмент мегантропа (Meganthropus paleojavanicus) Сангиран 6 (1) в сравнении с соответствующей частью нижней челюсти современного человека (Homo sapiens) (2), самца гориллы (Gorilla gorilla) (3) и самца орангутана (Pongo pygmaeus) (4).
На внутренней стороне челюсти рельеф выражен тоже нерезко: поверхность, служащая для отхождения подбородочно-язычной мышцы (m. genioglossus), сходна по положению с гоминидным типом. На ней находитсянебольшой острый выступ, соответствующий по месту расположения характерной для гоминин подбородочной ости (spina mentalis). Никакого образования, аналогичного "обезьяньей полке" (lamina simialis), на внутренней стороне симфиза не заметно. В подбородочной области человеческих челюстей, на границе нижней и внутренней поверхности, находится сравнительно плоская, продолговатая ямка для двубрюшной мышцы (fossa digastrica). У мегантропа она несколько смещена на лингвальную сторону и имеет треугольную форму; поверхность треугольника слегка вогнута, а у его вершины поднимается небольшой бугорок. Восходящая ветвь, насколько можно судить, отстояла от тела челюсти в сторону.
Зубная дуга мегантропа V-образно изогнута, скорее напоминая таковую гоминин, а не U-образную дугу человекообразных обезьян. Ее передняя часть укорочена и округлена. Альвеола клыка находится значительно медиальнее p1, тогда как у человекообразных ее положение лишь очень мало смещено по отношению к линии, продолжающей прямую заклыковых зубов.
Компьютерные модели нижнечелюстного фрагмента мегантропа (Meganthropus paleojavanicus) Сангиран 6 и его зубов.
Нижние резцы относительно небольшие. Клык обладает невысокой коронкой и не отличается массивностью, несколько выдается из общего ряда. По своим размерам он уступает клыку человекообразных обезьян и сравним с таковым Homo erectus. Щечные зубы, несмотря на значительные размеры, превышающие все известные зубы ископаемых людей и человекообразных обезьян, по своему строению вполне человеческие. Жевательные поверхности отличаются значительной изношенностью.
Строение коронки р1 не вполне симметричное. Она несет 2 бугорка, которые приблизительно одинаково развиты; обе "ямочки" на коронке отчетливо видны. Имеется 1 раздвоенный корень (что бывает и у гоминин). Почти по всем этим признакам р1 мегантропа резка отличается от соответствующего зуба человекообразных обезьян (по общей форме он секториальный; его продольная ось направлена косо относительно линии заклыковых зубов; обычно он имеет 2 корня и 1 сильно выступающий бугорок; средняя и задняя "ямочки" на жевательной поверхности его коронки не выражены).
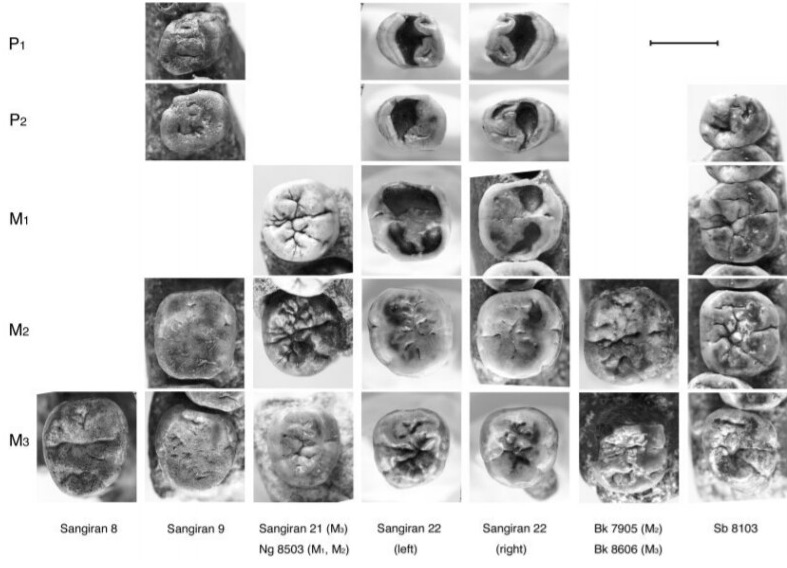
Нижнечелюстной фрагмент мегантропа (Meganthropus paleojavanicus) Сангиран 9 (слева) и нижняя челюсть человека выпрямленного (Homo erectus) Сангиран 22 (справа).
р2, будучи немного крупнее р1, отличается строением, еще более типичным для гоминин. Он также обладает единственным корнем. Двубугорковое строение коронки вполне ясно выражено, но коронка более асимметрична, чем на р1. Бугорки смещены на ее мезиальную сторону, оставаясь почти равными друг другу по размерам. Талонид этого зуба развит заметно сильнее тригонида. Передняя и задняя "ямочки", хотя и выражены, но не так отчетливо, как на р1. На язычной стороне коронки видны остатки цингулюма.
m1, кроме величины, вполне укладывается в рамки вариаций коренных зубов гоминин. На щечной стороне коронки хорошо выражены остатки пояска, на язычной — слабее. Форма коронки указывает на преобладание талонида. Число бугорков на жевательной поверхности достигает 5 или 6. Расположение бугорков и бороздок образует дриопитековый узор. m1 имеет 2 корня, из которых задний уклоняется в дистальном направлении. По-видимому, прорезался этот зуб очень рано, как это наблюдается у современного человека. m2 и m3 также двухкорневые, в то время как коренные Homo erectus имеют единственный корень.
Жевательные поверхности щечных зубов мегантропа (Meganthropus paleojavanicus) (Сангиран 8 и 9) и человека выпрямленного (Homo erectus).
Итак, для зубочелюстного материала мегантропа характерно сочетание архаичных (очень большая толщина нижней челюсти, отсутствие на ней подбородочного выступа, цингулюма на предкоренных) и прогрессивных, типичных для рода Homo признаков (короткость нижней челюсти и, очевидно, слабый прогнатизм, высокое положение подбородочного отверстия, наличие подбородочной ости, отсутствие "обезьяньей полки", положение ямки двубрюшного мускула, форма предкоренных зубов). Вследствие этого долго преобладала точка зрения, согласно которой мегантроп представляет собой специализированную форму древнего человека.
Поскольку характер износа коренных зубов соответствует таковому орангутанов, высказано предположение, что мегантроп питался фруктами и другими частями растений, растущими над землей, подобно современным орангутанам. У Homo erectus, вероятно, была более гибкая диета, благодаря его способности готовить пищу различными способами. Однако на сегодняшний день не существует никаких свидетельств в пользу того, что однообразная диета или даже сам человек прямоходящий каким-то образом способствовали исчезновению мегантропа.
Таким образом, около 1 млн. лет назад на территории современной Явы обитало по меньшей мере 4 вида гоминид — орангутан (Pongo sp.), гигантопитек Блэка (Gigantopithecus blacki), мегантроп (Meganthropus palaeojavanicus) и человек выпрямленный (Homo erectus).
Домыслы и спекуляции
Мегантроп нередко оказывается в центре различных спекуляций. Наверное, самое известное недоказанное предположение — то, что он был гигантом, имея по оценке Ф. Вейденрайха рост около 240 см и вес порядка 250 кг. Не подлежит сомнению, что для мегантропа характерны чрезвычайно крупные и массивные черепные кости, однако представляется весьма вероятным, что представление о его очень большом (более 200 см) росте может оказаться ошибочным. Костного материала для подтверждения подобных предположений имеется явно недостаточно, а тот, что есть в наличии, по мнению ряда исследователей, не превосходит по своим параметрам таковой Homo erectus. Рассчитанная Г. Крантцом на основе нижнечелюстного фрагмента Сангиран 6 высота черепа мегантропа составляет порядка 21 см, что соответствует росту менее 200 см.
Существуют неподтвержденные и весьма противоречивые слухи о посткраниальном материале, якобы принадлежащему мегантропу, но в серьезной научной прессе сообщения о них отсутствуют. Р. Гилрой приписывает мегантропу некие гигантские инструменты, будто бы обнаруженные в Австралии. Тем не менее, большинство палеоантропологов разделяет точку зрения, согласно которой мегантроп достоверно известен лишь с острова Ява.
Что же касается предположения о том, что мегантроп якобы дожил до наших дней и является т. н. "снежным человеком" (или йети, бигфутом и т. п.), а также того, что он будто бы послужил прообразом библейских гигантов, то их приходится признать ничем не подтвержденными фантазиями.
Ссылки
Meganthropus Skull (Bone Clones)
перейти
Литература
Дробышевский С. В. Предшественники. Предки? Архантропы. Гоминиды, переходные от архантропов к палеантропам. — М.: Едиториал УРСС, 2004
Елинек Я. Большой иллюстрированный атлас первобытного человека. — Прага: Артия, 1983
Зубов А. А. Палеоантропологическая родословная человека. — М.: Россельхозакадемия, 2004
Ископаемые гоминиды и происхождение человека / Отв. ред. Бунак В. В. — М.: Наука, 1966
Ларичев В. Е. Сад Эдема. — М.: Издательство политической литературы, 1980
Рогинский Я. Я., Левин М. Г. Антропология. Учебное пособие. — М.: Высшая школа, 1978
Эйдельман Н. Я. Ищу предка. — М.: Молодая гвардия, 1970
Якимов В. П. Гигантские ископаемые антропоиды (Hominidae?). Природа 12, 1947
Ciochon R. The mystery ape of Pleistocene Asia. Nature 459 (7249), 2009
Ciochon R., Olsen J., James J. Other Origins: The Search for the Giant Ape in Human Prehistory. — New York: Bantam Books, 1990
Durband A. C. A re-examination of purported "Meganthropus" cranial fragments. American Journal of Physical Anthropology 36, 2003
Encyclopedia of Human Evolution and Prehistory, 2nd ed. / Ed. by Delson Е., Tattersall I., Van Couvering J., Brooks A. S. — New York: Taylor & Francis, 2000
Kaifu Y., Aziz F., Baba H. Hominid Mandibular Remains From Sangiran: 1952-1986 collection. American Journal of Physical Anthropology 128 (3), 2005
Kaifu Y., Baba H., Aziz F., Indriati E., Schrenk F., Jacob T. Taxonomic affinities and evolutionary history of the Early Pleistocene hominids of Java: dentognathic evidence. American Journal of Physical Anthropology 128 (4), 2005
Koenigswald G. H. R. Australopithecus, Meganthropus and Ramapithecus. Journal of Human Evolution 2 (6), 1973
Kramer A. A critical analysis of claims for the existence of Southeast Asian australopithecines. Journal of Human Evolution 26 (1), 1994
Kramer A., Konigsberg L. W. The phyletic position of Sangiran 6 as determined by multivariate analysis. Courier Forschungs-institut Senckenberg 171, 1994
Marks P. Preliminary note on the discovery of a new jaw of Meganthropus von Koenigsvald in the lover Middle Pleistocene of Sangiran, Central Java. Indonesian Journal of Natural Science 1, 2, 3, 1953
Orban-Segebarth R., Procureur F. Tooth size of Meganthropus palaeojavanicus. Journal of Human Evolution 12 (8), 1983
Paleoanthropology, Morphology and Paleoecology / Ed. by Tuttle R. H. — La Hague: Mouton, 1975
Robinson J.T. Further remarks on the relationship between "Meganthropus" and Australopithecines. American Journal of Physical Anthropology 13 (3), 1955
Robinson J. T. Meganthropus, australopithecines and hominids. American Journal of Physical Anthropology 11 (1), 1953
Sartono S., Tyler D. E., Krantz G. S. A new 'Meganthropus' mandible from Sangiran, Java: an announcement. Human Evolution in Its Ecological Context 1, 1995
Sémah F., Sémah A., Djubiantono T. They Discovered Java. — Bandung: Pusat Penelitian Arkeologi Najional, 1990
Szalay F., Delson E. Evolutionary history of the Primates. — New York: Academic Press, 1979
The Primate Fossil Record / Ed. by Hartwig W. C. — Cambridge: Cambridge University Press, 2002
Tyler D. Е. A Taxonomy of Javan hominid mandibles. Human Evolution 6, 1991