Мегистотерии — Megistotherium
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Клада Скротиферы (Scrotifera)
Клада Феревунгуляты (Fereuungulata)
Клада Ферэ (Ferae)
Клада Всехищные (Pan-Carnivora)
Отряд †Гиенодонты (Hyaenodonta)
Надсемейство †Гиайнайлюроиды (Hyainailouroidea)
Семейство †Гиайнайлюровые (Hyainailouridae)
Подсемейство †Гиайнайлюрины (Hyainailourinae)
Род †Мегистотерии (Megistotherium)
Вид †Megistotherium osteothlastes
Мегистотерии (Megistotherium Savage, 1973) — монотипический род подсемейства гиайнайлюрин (Hyainailourinae (Pilgrim, 1932)), происходящий из раннего - среднего миоцена Африки. Единственный установленный вид — Megistotherium osteothlastes Savage, 1973 (тип).

Реконструкция внешнего облика мегистотерия (Megistotherium osteothlastes). Автор — Р. Евсеев.
Этимология
Название рода и вида дословно переводится с латыни как "огромный зверь-костедробитель".
Находки
Наиболее полные ископаемые остатки мегистотерия были обнаружены в 1961-1966 гг. в раннемиоценовых (аквитанский / бурдигальский ярус, 23-16 млн. лет назад) отложений формации Марадаг из Джебель-Зелтен в Ливии. Эти находки представлены довольно хорошо сохранившимся черепом не старого еще животного с отсутствующими зубами, 4 осколками другого черепа и 9 посткраниальными фрагментами (атлант, дистальные фрагменты плеча, астрагал, метаподии и др.). На основании данного материала был описан типовой и пока единственный четко установленный вид мегистотериев — Megistotherium оsteothlastes.



Череп мегистотерия (Megistotherium osteothlastes) из Джебель-Зелтен, Ливия.
В раннемиоценовых (MN 4, бурдигальский ярус, 17-16 млн. лет назад) слоях формации Могхара на севере Египта был найден правый фрагмент нижней челюсти с p4-m3, а также небольшой, но относительно предыдущего более крупный нижнечелюстной фрагмент с m1-2, принадлежащие Megistotherium osteothlastes.
Средним миоценом (серравалльский ярус, от 13 до 12 млн. лет назад) датируются правый m2 и дистальный фрагмент левого плеча Megistotherium osteothlastes, происходящие из местонахождений Бартуле и Кабарсеро формации Нгорора (Туген-Хиллз, район Баринго, Кения). Несколько старшим возрастом (14,5 млн. лет назад) датирован левый верхний клык из Чепараво, также в Туген-Хиллз.

Атлант мегистотерия (Megistotherium osteothlastes) из Джебель-Зелтен, Ливия.
Небольшое количество разрозненных скелетных остатков, также причисляемых к Megistotherium оsteothlastes, извлечено из отложений Форт-Тернана в Центральной Ньянзе, Кения, имеющих среднемиоценовый возраст (серравалльский ярус, 13,7 млн. лет назад).
Другие восточноафриканские находки известны из местонахождений Рузинга, Мабоко, Омбо и Кипсараман на территории Кении, а также Морото I в Уганде.

Фрагмент правой ветви нижней челюсти мегистотерия (Megistotherium osteothlastes) из Египта.
Помимо вышеперечисленного, имеется информация о плохо сохранившейся нижней челюсти с р2-3 из слоев Бугти в Пакистане, а также разрозненных крупных зубах из первой половины миоцена Белуджистана, Пакистан. Принадлежность этих остатков Megistotherium весьма спорна и устанавливается главным образом из-за их значительных размеров. Гораздо более вероятно, что на самом деле они принадлежат одному из видов гиайнайлюров — Hyainailouros bugtiensis.

Остатки мегистотерия (Megistotherium оsteothlastes) из Туген-Хиллз, Кения: слева — левый верхний клык, снаружи и изнутри; справа — m2 (снаружи, сверху и изнутри), внизу — дистальный фрагмент левого плеча.
Строение и функции
Имеющиеся остатки посткрания мегистотерия настолько скудны, что это значительно ограничивает наши представления о его внешнем облике. Установлено, что этот гиенодонт имел крупную голову, удлиненное приземистое туловище и короткие мощные конечности. Затруднительным оказывается также вычисление его общих размеров, которое пока можно произвести лишь предположительно. По приблизительным подсчетам, длина тела с головой составляла 3,8-4 м (не учитывая длину хвоста, который, возможно, имел около 1,5 в длину), а рост в холке достигал порядка 1,9 м.
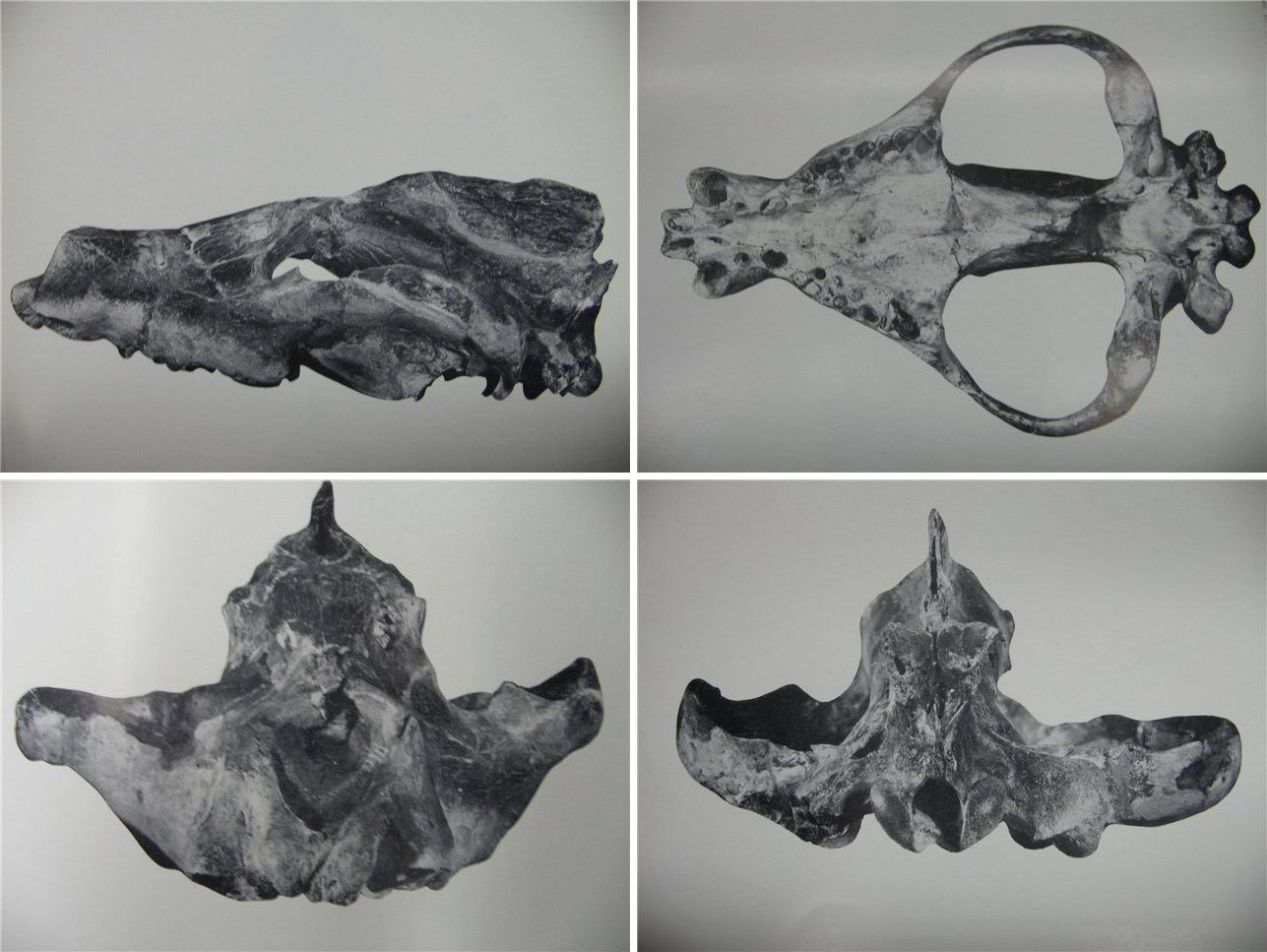
Череп мегистотерия (Megistotherium osteothlastes) из Джебель-Зелтен, Ливия.
Наиболее полная и хорошо изученная находка — череп без нижней челюсти, обнаруженный в Ливии. Его длина составляет 66,4 см, а ширина в области височных дуг достигает 47,1 см. Исходя из размеров найденного позже фрагмента нижней челюсти, а также учитывая принадлежность ливийского черепа не старой, а значит и не самой крупной особи, было заключено, что череп мегистротерия мог достигать и гораздо большей величины — около 90 см в длину. При подобных размерах масса этого хищника достигала 800-1000 кг, что ставит его в ряд наиболее крупных плотоядных млекопитающих. С другой стороны, найденная в Ливии часть плечевой кости по своему размеру вполне сравнима с медвежьей. Поэтому до появления новых находок нельзя исключать того, что обладая действительно огромной головой, мегистотерий имели гораздо более скромное по величине туловище (подобные пропорции были довольно характерны для крупных гиенодонтов). Так или иначе, даже самые осторожные подсчеты позволяют установить для этого хищника длину тела в 2,5-3 м (без учета хвоста) и вес около 400 кг. Помимо крупных размеров, мегистотерий отличается целым рядом своеобразных черт.

Сравнение черепов мегистотерия (Megistotherium osteothlastes), льва (Panthera leo), бурого медведя (Ursus arctos) и пятнистой гиены (Crocuta crocuta).
Одним из важнейших биологических признаков, лежащих в основе стратегии поведения и адаптивных возможностей каждого вида, является ориентация глазниц, или глазных осей: латеральная, фронтальная или промежуточная. У представителей подсемейства Hyaenodontinae наблюдается почти предельный латеральный тип ориентации глазниц (угол между осью глазницы и сагиттальной плоскостью черепа приближался к 90°), обеспечивая максимальный угол обзора, что характерно для обитателей открытых пространств. Это сильно отличает их от других вымерших и современных хищников. А у Hyainailourinae, и мегистотерия в том числе, по всей видимости, был промежуточный — латеромедиальный тип ориентации глазниц, что конвергентно сближает их с представителями семейств псовых (Canidae) и кошачьих (Felidae). Подобная направленность глазниц характерна для наземных специализированных хищников с широким сектором бинокулярного зрения.
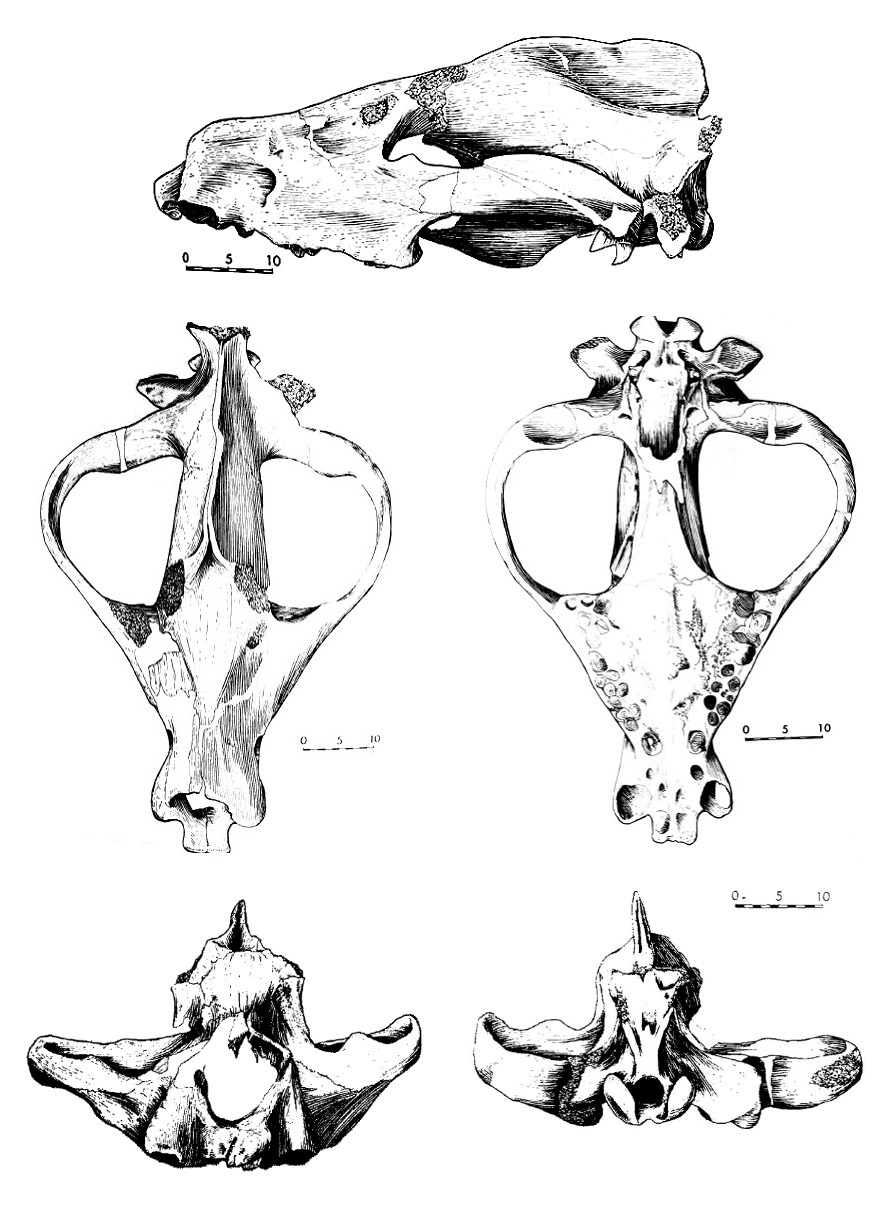
Прорисовки черепа мегистотерия (Megistotherium osteothlastes) из Джебель-Зелтен, Ливия.
Череп мегистотерия, как и у большинства Hyainailourinae, имеет широко расставленные полукруглые (при виде сверху) скуловые дуги. Их весьма значительная ширина, а также очень мощный сагиттальный гребень свидетельствуют об исключительном развитии височных мышц (масса каждой из них оценивается приблизительно в 10 кг). В то же время скуловые дуги гораздо менее массивны, чем этого можно было бы ожидать при таких их размерах. Это означает, что жевательные мышцы не были развиты в той же степени, что и височные, и последние играли основную роль при закрывании челюстей, как и у большинства хищных (Carnivora). Строение челюстного сустава, в целом сходное с таковым у кошачьих (Felidae), обеспечивало открывание нижней челюсти под углом в 50-60°. В таком случае, при отведении челюсти расстояние между вершинами клыков составляло около 30 см. Лицевой отдел черепа мегистотерия довольно длинный, что, помимо прочего, можно объяснить и особенностями их рациона.
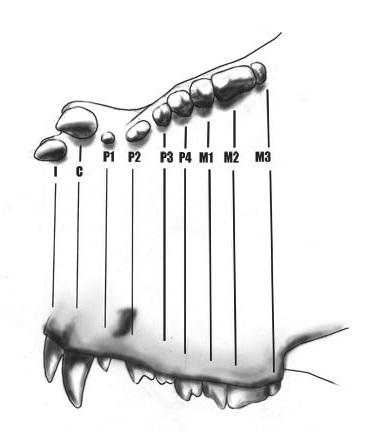
Прорисовки реконструированного верхнего зубного ряда мегистотерия (Megistotherium osteothlastes).
Зубная формула: I1/i3, C1/c1, P4/p4, M3/m3. В отличие от большинства Hyainailourinae, в верхней челюсти имеется только 1 пара резцов, а не 3. Сохранившиеся резцы (вероятно, I3) крупные и мощные, функционально дополняющие клыки. Сокращение числа резцов и специализация при этом одной из пар уменьшают возможность их обламывания. Из современных хищников, кстати, I3 наиболее увеличены у крупных кошачьих (Felidae) и гиеновых (Hyaenidae). К сожалению, достоверные находки клыков мегистотерия неизвестны, однако сохранившиеся альвеолы указывают на то, что они относительно других зубов были более смещены наружу, имели крупные размеры и были овальными в сечении. Впрочем, найденные в Кении разрозненные клыки обладают подходящими для этого гиенодонта размерами и формой, так что вполне могут ему принадлежать. Если челюсти рассматривать в качестве рычага, уровень эффективности использования клыков зависит от их размера и близости их расположения к месту крепления щечной мускулатуры. Следовательно, укороченная лицевая часть черепа пятнистой гиены (Crocuta crocuta) и большой панды (Ailuropoda melanoleuca) существенно повышает эффективность действия их клыков.
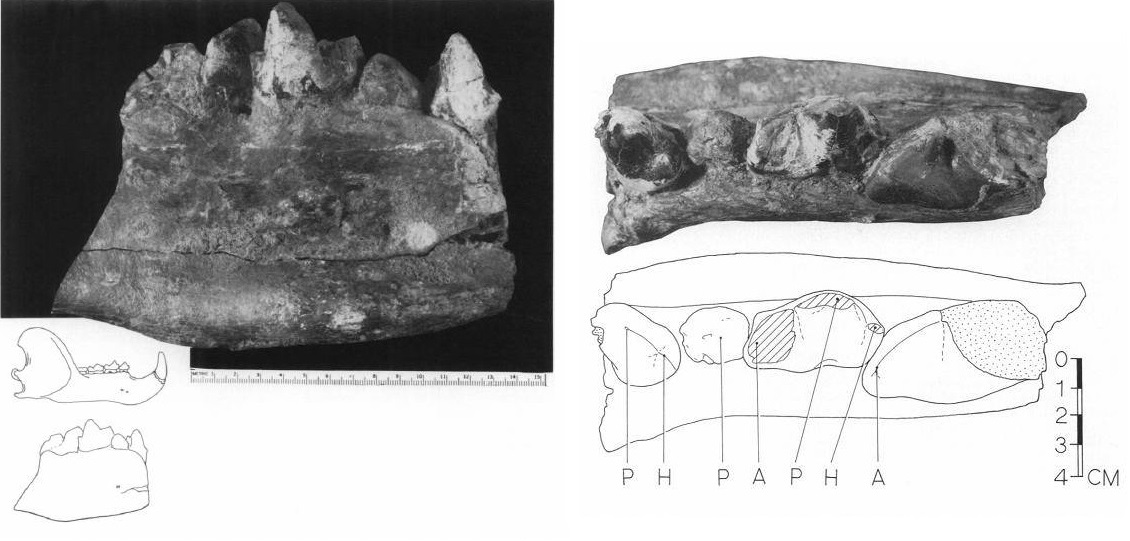
Фрагмент правой ветви нижней челюсти мегистотерия (Megistotherium osteothlastes) с p4-m3 из отложений формации Мохара, Египет, снаружи и сверху. В вставке слева внизу он показан в сравнении с нижней челюстью белого медведя (Ursus maritimus) — крупнейшего современного наземного хищника. На прорисовке жевательной поверхности справа отмечены: А — параконид, Н — гипоконид, Р — протоконид.
Наоборот, отличительной особенностью черепа мегистотерия является удлиненный лицевой отдел. Однако даже если принять во внимание этот признак недостатка силы укуса, необходимо учитывать, что отношение площади клыка к его расстоянию до места крепления щечной мускулатуры у мегистотерия больше, чем у всех современных карнивор. Это предполагает, что клыки являлись основным орудием добывания пищи для этого гигантского хищника. Р1 (как и Р2) имеет 1 корень и располагается сзади и сбоку от клыка, в месте наибольшего сужения неба. Предкоренные массивны и обладают затупленными коронками. Невозможно утверждать наверняка, однако представляется весьма вероятным, что они действовали не менее эффективно, чем у современных гиен, выдерживая значительное давление при сжатии мощных челюстей. Р3-4 мощные, с 3 корнями; ширина Р4 превышает его длину; М1-2 острые; М3 небольшой и расположен поперечно.
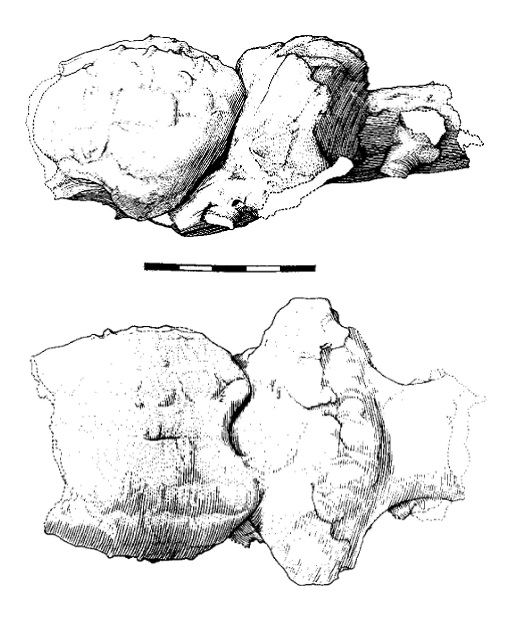
Прорисовки слепка головного мозга мегистотерия (Megistotherium оsteothlastes), по черепу из Джебель-Зелтен, Ливия.
Нижняя челюсть высокая, с массивным внешним валиком по своему нижнему краю. Передний отдел нижней челюсти не сохранился, однако можно предположить, что он нес 2 функциональные и 1 рудиментарную пару резцов (согласно иной точке зрения, все нижние резцы были сильно редуцированны и при жизни скрывались в мягких тканях десны). Вероятно, нижние клыки также имели крупные размеры. p4 примечателен смещенной ориентацией относительно оси нижней челюсти и отличается от такового у Hyainailouros маленьким, острым гипоконидом. m1 отличается от таковых у Pterodon и Hyaenodon чрезвычайной редукцией и отсутствием дифференцированных острых выступов и гребней, его единственный цилиндрический корень достигает большего диаметра, чем коронка. m2-3 напоминают таковые у большинства Hyainailouridae, но в отличии от Apterodon обладают длинным лезвиеподобным параконидом и (по крайней мере, m2) маленьким гипоконидом. По своему строению m3 напоминает двухкорневой m2, но существенно превосходит его по размерам. Длина коронки этого зуба достигае 7-8 см, что делает m3 мегистотерия самым большим известным коренным зубом хищного млекопитающего. Сохранившиеся нижние щечные зубы несут явные следы износа. Непосредственно под m1 находится относительно небольшое нижнечелюстное отверстие.
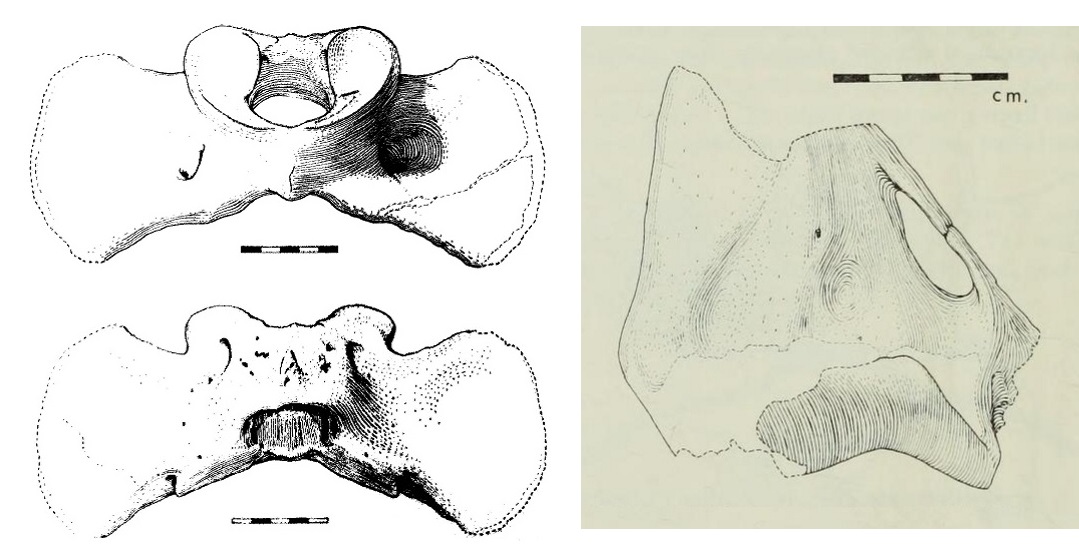
Слева: прорисовки атланта мегистотерия (Megistotherium osteothlastes) из Джебель-Зелтен, Ливия; сверху и снизу.
Справа: прорисовка дистального конца правой плечевой кости мегистотерия (Megistotherium osteothlastes) из Джебель-Зелтен, Ливия; спереди.
Строение атланта и боковых затылочных костей мегистотерия свидетельствует о сильном развитии мышц, поворачивающих голову в стороны. Видимо, они могли быть активно задействованы и при отрывании плоти при питании, а также при преодолении сопротивления добычи. Судя по строению зубной системы — выдерживающим большую нагрузку усиленным верхним клыкам и дополняющим их верхним резцам, — мегистотерий мог наносить мощнейшие укусы. Судя по промежуткам между верхними резцами и клыками, нижние клыки прикусывали перед верхними, а чрезвычайно развитые височные мышцы свидетельствуют об огромной силе сжатия челюстей. Представляется вероятным, что этот гиенодонт был способен намертво вцепиться в жертву и вырвать целый кусок сильным движением головы из настигнутой добычи или уже повергнутой — при поедании. При этом он наверняка помогал себе мощными конечностями.

Этапы портретной реконструкции мегистотерия (Megistotherium osteothlastes). Материалом послужили череп из Ливии, нижнечелюстной фрагмент из Египта, образцы зубов Hyainailouros и находки нижних челюстей и зубов Pterodon. Верхние зубы подогнаны под альвеолы. Ливийский череп подогнан под размер фрагмента нижней челюсти в соответствии с окклюзией зубов. Автор — Р. Евсеев.
Эволюционные связи
Предки мегистотерия — гиенодонты из подсемейства гиайнайлюрин (Hyainailourinae) — известны в Африке начиная с позднего эоцена. Собственно род Megistotherium возник в начале миоцена, когда эпоха господства гиенодонтов уже близилась к своему завершению, и венчает собой африканскую ветвь их развития. Находки из Кении свидетельствуют о том, что мегистотерий просуществовал по меньшей мере до конца среднего миоцена (11,6 млн. лет назад).

Портретная реконструкция мегистотерия (Megistotherium osteothlastes). Автор — Р. Евсеев.
Ближайшими родственниками мегистотериев считаются птеродоны (Pterodon) и очень крупные гиайнайлюры (Hyainailouros), достаточно широко распространенные в Старом Свете. На основании анатомических, хронологических и географических данных высказано предположение, что очень схожие по строению миоценовые Megistotherium и Hyainailouros происходят от общего предка, ведущего начало от одного из крупных представителей олигоценового рода Pterodon, например Pterodon africanus из Файюма в Египте. В то же время необходимо учитывать, что установленные морфологические различия Hyainailouros и Megistotherium довольно незначительны и выражаются у последнего главным образом в косой ориентации р4, присутствии на нем гипоконида и более крупных размерах тела. По мнению ряда исследователей, упомянутых черт явно недостаточно для разделения этих 2 родов, однако пересматривать их таксономию следует лишь после обнаружения более полного ископаемого материала (особенно это касается Hyainailouros), анализ которого предоставит возможность установить более весомые диагностические признаки.

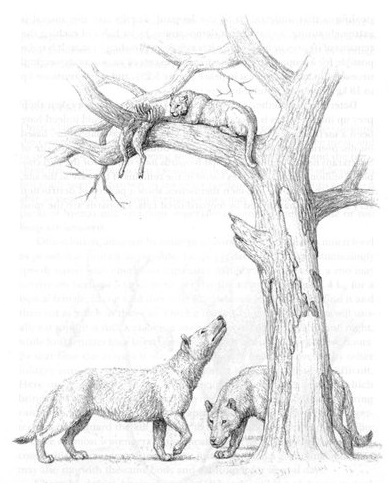
Слева: мегистотерий (Megistotherium оsteothlastes) в естественном окружении. Справа: пара мегистотериев (Megistotherium оsteothlastes), над ними на дереве — афросмил (Afrosmilus africanus) с добычей (по находкам из Джебель-Зелтен, ранний миоцен Ливии). Автор — М. Антон.
Образ жизни
Таким образом, имеющиеся данные свидетельствуют в пользу того, что мегистотерий специализировался на добыче крупнейших фитофагов своего времени, с их толстой шкурой и мощным костяком — и был способен справляться как с тем, так и с другим. Помимо активной охоты на крупную добычу, этот огромный гиенодонт, вероятно, был неплохо приспособлены и к поеданию падали (судя по строению предкоренных). Вероятно, открытая пасть мегистотерия была достаточно широкой, чтобы охватить ногу современных ему хоботных. По всей видимости, в то время в Африке просто не существовало других хищников, способных совладать как с остатками, так и с живой добычей такого размерного класса. Следует отметить, правда, что близкий по размерам и строению род Hyainailouros распространился куда шире мегистотерия. Тем не менее, не будучи многочисленным, последний просуществовал в Африке более 10 млн. лет. Настоящие гиены-некрофаги, способные перемалывать кости, в те времена еще не появились, однако к концу олигоцена на Африканском континенте обособилась группа специализированных гиенодонтов-склерофагов из подсемейства Teratodontinae.

Мегистотерий (Megistotherium osteothlastes) атакует гомфотерия (Gomphotherium angustidens). Автор — Р. Евсеев.
Сопутствующие находки свидетельствуют, что рядом с мегистотерием жили антракотерии (Masritherium, Brachyodus), бегемоты (Kenyapotamus), свиньи (Bunolistriodon, Listriodon), ранние носороги (Brachyodus, Paradiceros, Aceratherium), примитивные хоботные (Prodeinotherium, Gomphotherium, Zygolophodon) и др. Многие из этих животных были адаптированы к потреблению древесно-кустарниковой и водной мягкой растительности, обитая в околоводных и лесистых районах, предоставляющих им обильную пищу. Тех же мест обитания наверняка придерживался и охотившийся на них мегистотерий. Находки ископаемых остатков этого гиенодонта относительно редки, что указывает на небольшую плотность его популяции и косвенно подтверждает хищный образ жизни — таких больших хищников попросту не может быть много, иначе для них не хватит пищи.

Мегистотерий (Megistotherium osteothlastes) прогоняет перкрокут (Percrocuta sp.). Автор — Р. Евсеев.
Исчезновение
Начиная примерно с середины миоцена климат Земли начинает меняться, все более приближаясь к современному. Постепенное снижение среднегодовых температур привело к уменьшению площади влажных биотопов и замещению их травянистыми степями. Все большее распространение получают конкурирующие с гиенодонтами и более пластичные в эволюционном смысле настоящие хищные (Carnivora). Приспосабливаясь к нарастающему давлению с их стороны, бурно развиваются растительноядные млекопитающие. Широкое распространение получают такие их быстроходные формы, как лошади и антилопы. Совокупность всех этих факторов и привела последних гиенодонтов к вымиранию. На примере мегистотерия мы видим, как увеличивая размеры и массу, гиенодонты сохраняли при этом примитивный уровень организации. Очевидно, именно подобная консервативность и стала для них роковой.
Ссылки
Megistotherium (Paleobiology Database) перейти
Литература
Лавров А. В. Неизвестные хищники креодонты. ПалеоМир 1 (8), 2010
Савельвев С. В., Лавров А. В. Окаменевшие мозги. Химия и жизнь 1, 2002
Bishop W. W., Miller J. A., Fitch F. J. New Potassium-Argon Age Determinations Relevant to the Miocene Fossil Mammal Sequence in East Africa. American Journal of Science 267 (6), 1969
Ginsburg L. Hyainailouros sulzeri, mammifère créodonte du Miocène d’Europe. Annales de Paléontologie 66, 1980
Morales J., Pickford M. Carnivores from the Middle Miocene Ngorora Formation (13-12 ma), Kenya. Estudios Geológicos 61 (3-6), 2005
Morales J., Pickford M. Creodonts and carnivores from the Middle Miocene Muruyur Formation at Kipsaraman and Cheparawa, Baringo District, Kenya. Comptes Rendus Palevol 7 (8), 2008
Morales J., Pickford M., Fraile S., Salesa M.J., Soria D. Creodonta and Carnivora from Arrisdrift, early Middle Miocene of southern Namibia. Memoir of the Geological Survey of Namibia 19, 2003
Morlo M., Miller E. R., El-Barkooky A. N. Creodonta and Carnivora from Wadi Moghra, Egypt. Journal of Vertebrate Paleontology 27, 2007
Naoko Е. Body Mass Estimates in Extinct Mammals from Limb Bone Dimensions: the Case of North American Hyaenodontids. Palaeontology 44 (3), 2001
Rasmussen D.T., Christopher D.T., Simons E.L. New Specimens of the Giant Creodont Megistotherium (Hyaenodontidae) from Moghara, Egypt. Journal of Mammalogy 70 (2), 1989
Savage R. J. G. Megistotherium, gigantic hyaenodont from Miocene of Gebel Zelten, Libya. Bulletin of the British Museum (Natural History), Geology 22 (7), 1973
Solé F., Lhuillier J., Adaci M., Bensalah M., Mahboubi M., Tabuce R. The hyaenodontidans from the Gour Lazib area (?Early Eocene, Algeria): implications concerning the systematics and the origin of the Hyainailourinae and Teratodontinae. Journal of Systematic Palaeontology 12 (3), 2014
Sorkin B. A biomechanical constraint on body mass in terrestrial mammalian predators. Lethaia 41 (4), 2008
Turner А., Antоn М. Evolving Eden: an illustrated guide to the evolution of the African large-mammal fauna. — New York: Columbia University Press, 2004