Многобугорчатые — Multituberculata
Клада Млекопитающеобразные (Mammaliaformes)
Клада †Аллотерии (Allotheria)
Отряд †Многобугорчатые (Multituberculata)
Многобугорчатые, или мультитуберкуляты (Multituberculata Cope, 1884), — отряд клады аллотериев (Allotheria Marsh, 1880), происходящий из средней юры - позднего палеоцена Европы, поздней юры - позднего эоцена Северной Америки, ? ранней / средней юры - раннего эоцена Азии, раннего мела Австралии и позднего мела Мадагаскара. Таксономически многобугорчатые весьма разнообразны: в их рамках установлены 2 подотряда, 6 надсемейств и 21 семейство. Это наиболее долго существующий и самый многочисленный отряд мезозойских млекопитающеобразных.
Многобугорчатое в естественном окружении.
Свое название группа получила в связи с характерным строением коренных зубов: их окклюзионные поверхности несут многочисленные бугорки (бугорок на латинском языке — tuberculum), расположенные рядами.
По величине многобугорчатые варьируют от мелкой мыши до бобра, однако большинство из них имеют размеры средней крысы.
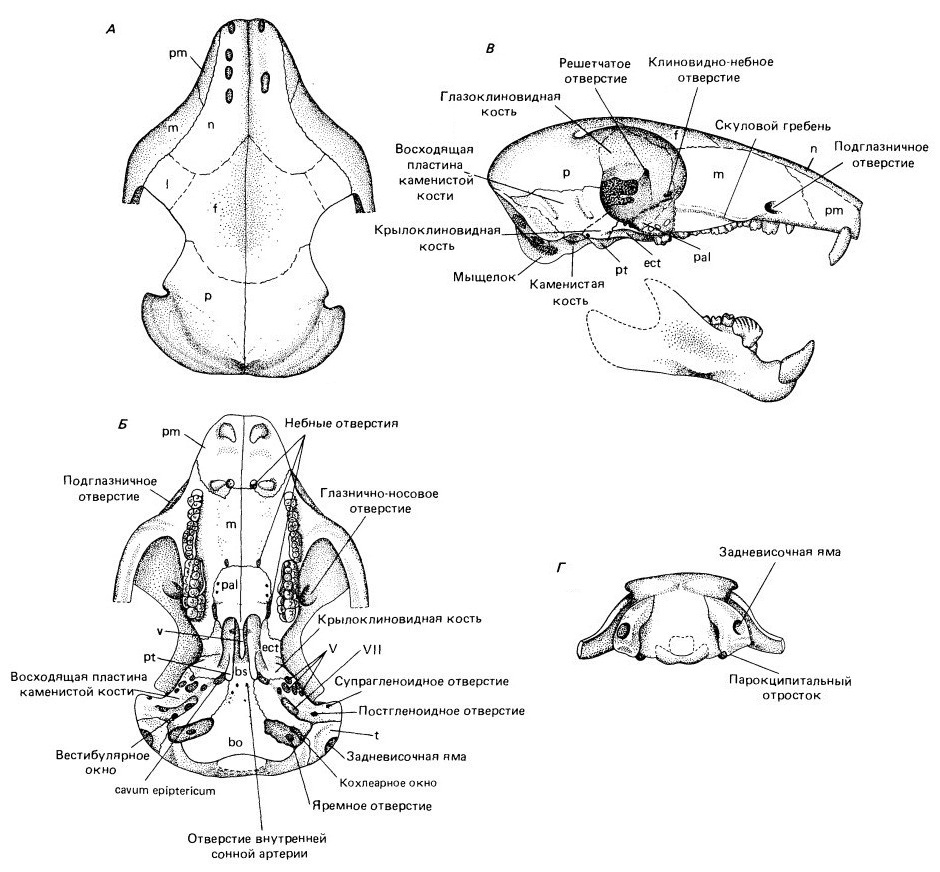
Прорисовки частично реконструированных черепа и нижней челюсти камптобаатара (Kamptobaatar kuczynskii): сверху (А), со стороны неба (Б), сбоку (В) и сзади (Г) (затылок изображен в меньшем масштабе).
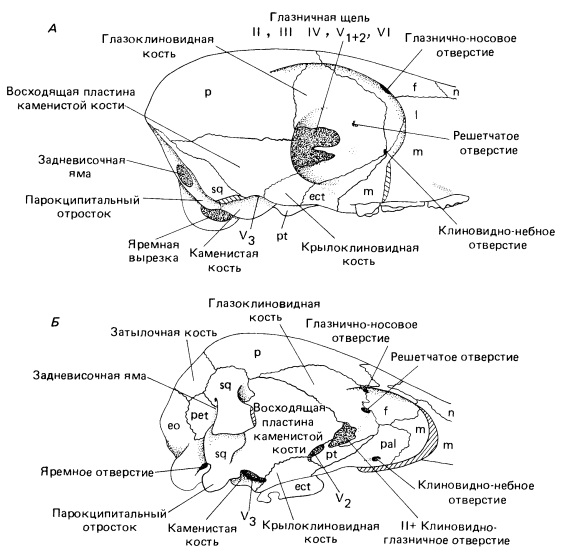
Череп многобугорчатых довольно крупный относительно остального тела. Он низкий и широкий, особенно в межглазничном пространстве. Лицевой отдел черепа короткий. Носовые кости большие, расширяющиеся назад. Лобные кости нависают над глазницами. Судя по устройству последних, глаза смотрели не вперед, как это характерно для большинства ранних териевых млекопитающих, а вбок. Скуловая дуга сформирована длинным задним отростком верхнечелюстной кости и чешуйчатой костью, сама же скуловая кость отсутствует. Мозговая коробка относительно мелкой крыловидной костью и крупной восходящей пластиной каменистой напоминает известную для морганукодонтов и триконодонтов. Чешуйчатая кость в образовании ее стенки существенно не участвует. Мозговая полость очень маленькая. Затылок примитивного типа, поскольку сохраняет задневисочную яму.
Прорисовки мозговой коробки камптобаатара (Kamptobaatar kuczynskii) (А) и утконоса (Ornithorhynchus anatinus) (Б).
Нижняя челюсть короткая и массивная. Срастание ее ветвей в симфизе непрочное, что свидетельствует о возможности односторонних движений каждой из ветвей челюсти. Передний отдел впереди заднего премоляра большей частью сильно понижен. Массетерная ямка крупная. Угловой отросток отсутствует, венечный отросток обычно низкий. У большинства родов ямка челюстного сустава открывается вперед. Ее строение позволяло нижней челюсти совершать не только вертикальные, но и продольные движения, тогда как поперечные "запрещены". Челюсть двигалась спереди назад (палинальный ход), что не встречается у современных млекопитающих, но является отличительной чертой аллотериев. Во время смыкания челюстей укус усиливался оттягиванием нижней челюсти назад (примерная аналогия с жеванием у дицинодонтов).
Подобно настоящим млекопитающим, но независимо от них, квадратная и сочленовная кости вошли в состав среднего уха, превратившись в молоточек и наковальню. Как и у других примитивных млекопитающеобразных, слухового барабана еще нет, а улитка не свернута спиралью. Головной мозг небольшой и примитивно устроенный.
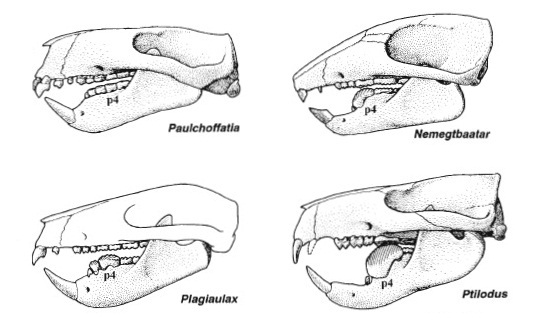
Прорисовки реконструированных черепов с нижними челюстями некоторых многобугорчатых с помеченным последним нижним предкоренным зубом (р4).
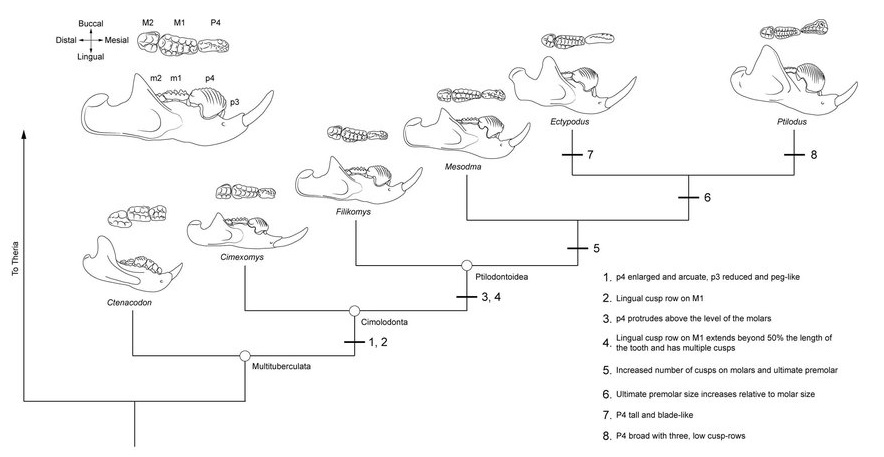
Зубная система сильно специализирована и напоминает таковую грызунов. Формула зубов: I2-3/i1, C1-0/c0, P1-5/p0-4, M2/m2-3. У наиболее архаичных форм зубы мало дифференцированы по величине, могут наличествовать верхние клыки, а щечные зубы сохраняют простое строение с многочисленными бугорками. Со временем число резцов редуцируется, они увеличиваются в размерах и приобретают постоянный рост. У типичных форм сохранившаяся II-я пара верхних резцов (I2) увеличена, направлена вниз; II-я пара нижних (i2) — еще более крупная, длинная, направлена вперед и несколько вверх (сходны с резцами грызунов), остальные отсутствуют. Клыки исчезают, а резцы отделяются от щечных зубов широкой диастемой.
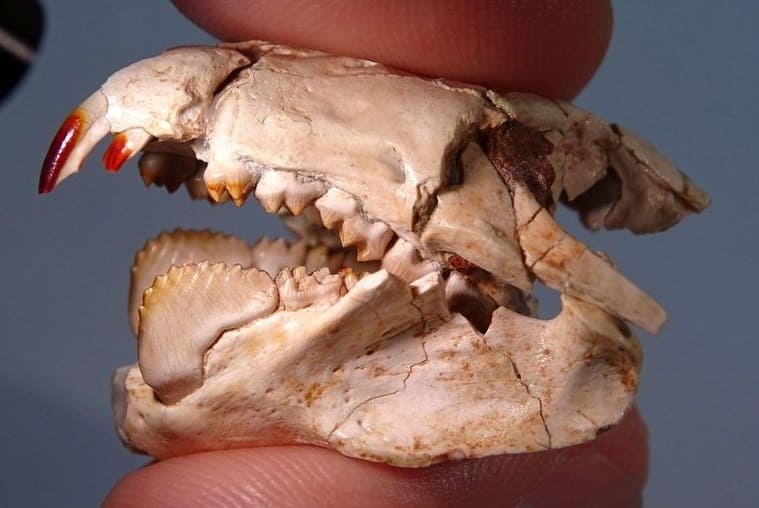
У представителей семейства Kogaionidae и надсемейства Taeniolabidoidea эмаль на передней поверхности резцов приобретает насыщенный красный цвет. Эта особенность также наблюдается у ряда ныне живущих землероек и грызунов и вызвана наличием в эмали минералов железа, которые, как считается, придают ей дополнительную прочность. В связи с разрозненностью материала неясно, унаследована ли эта морфологическая черта указанными группами многобугорчатых от общего предка, или же она развилась у них независимо.
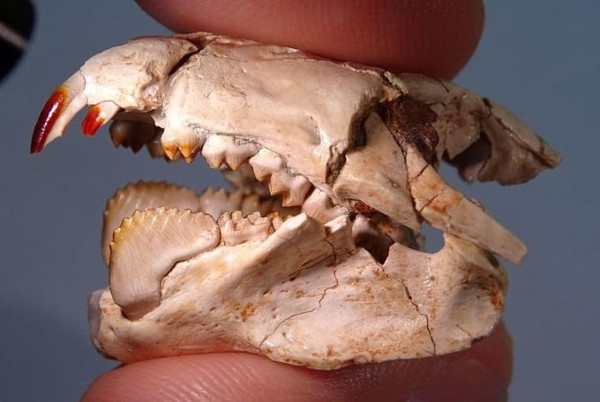
Неполный череп с нижней челюстью барбатодона (Barbatodon transylvanicus). Обращает на себя внимание укрепленная эмаль верхних резцов насыщенно-красного цвета.
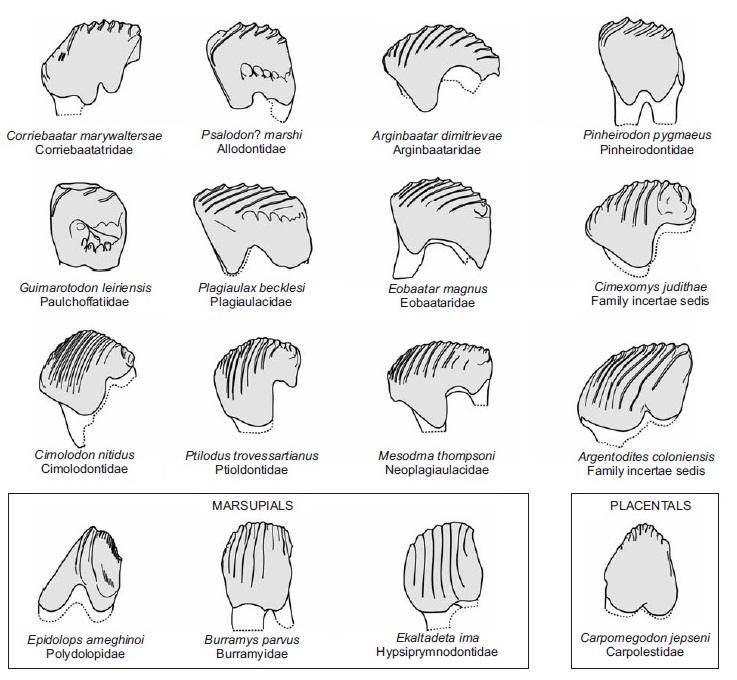
Щечные зубы режуще-перетирающего типа. Верхние предкоренные округлые или овальные, с небольшим числом бугорков, без режущего лезвия; P4 иногда усложнен и подобен коренным. Задний нижний предкоренной (p4) зачастую сильно удлиняется, становится значительно выше и существенно сжимается с боков, формируя продольное режущее срединное лезвие, зубчатое на вершине и с вертикально бороздчатыми боковыми поверхностями. Лезвиеподобные предкоренные являются апоморфией многобугорчатых; нечто подобное также наблюдается у некоторых кенгуру (например, у Hypsiprymnodontidae) и плезиадаписообразных Carpolestidae. Передние предкоренные значительно меньше, порой также со срединным режущим лезвием, продолжающим лезвие p4, или иногда, кроме рудиментарного p3, отсутствуют.
Прорисовки последнего нижнего предкоренного зуба (р4) различных зубов различных многобугорчатых, а также некоторых сумчатых и плацентарных.
Коренные зубы, по 2 наверху и внизу, длинные, с продольными рядами многочисленных бугорков (2-3 ряда на верхних коренных и 2 ряда на нижних), разделенными продольными бороздами. Предполагается, что дополнительные ряды бугров развились из бугорков цингулюма и цингугида. Бугорки имеют различную, иногда более или менее отчетливую поперечно-полулунную форму. При смыкании челюстей внешний ряд бугорков на нижних зубах входил в пространство между рядами на верхних. Строение, ориентация и характер износа коренных зубов подтверждает, что движение нижней челюсти происходило только в сагиттальной плоскости без какой-либо поперечной составляющей, присутствовавшей у наиболее примитивных млекопитающих.
Посткраниальный скелет многобугорчатых довольно однообразен и вполне соответствует генерализованному плану строения млекопитающеобразных. Туловище вытянутое и довольно приземистое. Таз узкий, с предлобковыми костями. Конечности пятипалые, стопоходящие. Хвост длинный. У форм, приспособленных к лазанью по деревьям, стопа приобрела способность разворачиваться назад, а хвост стал хватательным.
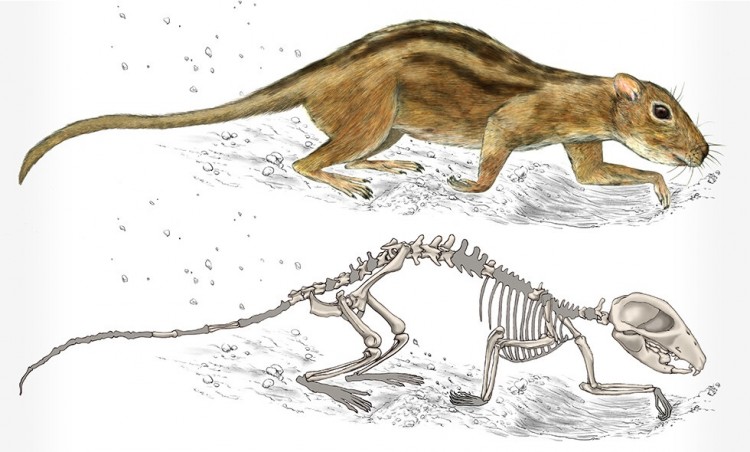
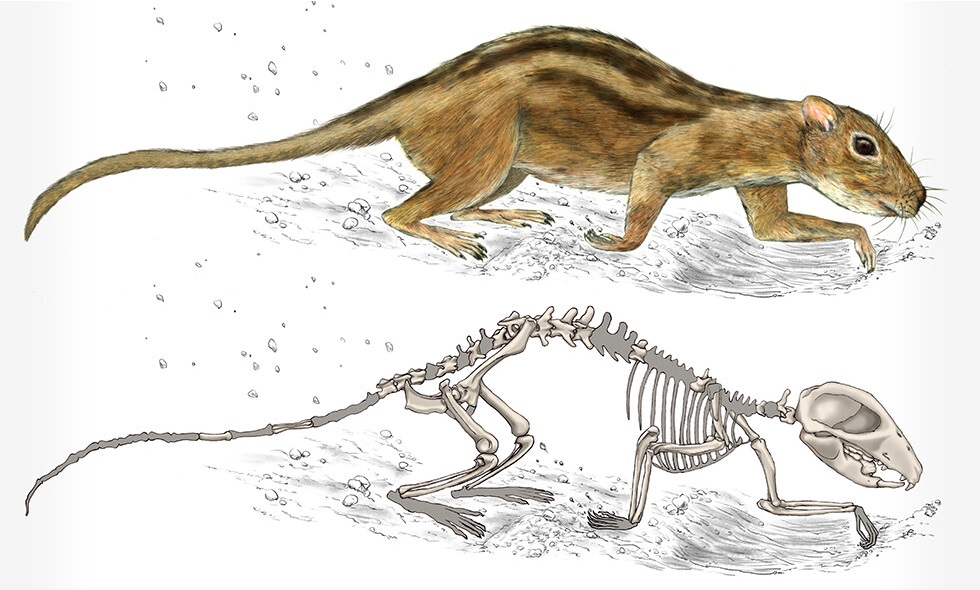
Прорисовка реконструированного скелета и реконструкция внешнего облика филикомиса (
Filikomys primaevus), устраивавшего подземные норы и жившего в них небольшими группами.
Современные живородящие млекопитающие используют две альтернативные стратегии размножения. Плацентарные отличаются долгим сроком беременности, но зато производят на свет крупных детенышей, которые довольно быстро становятся самостоятельными. Наоборот, у сумчатых беременность очень короткая, а детеныш рождается недоразвитым и вынужден ''дозревать'' в сумке своей матери. Хотя остатки детенышей многобугорчатых пока что не обнаружены, очень узкое пространство между половинками таза долгое время заставляло предполагать, что они, подобно современным сумчатым, рождали очень мелких детенышей, которые завершали свое развитие уже вне тела матери — может, и не в сумке, но по крайней мере в безопасном логове. На основании этого ''сумчатый'' способ размножения рассматривался как более древний, появившийся еще у общего предка териев и многобугорчатых, тогда как ''плацентарный'' развился уже у собственно плацентарных. Однако устоявшаяся точка зрения может пошатнуться благодаря новому исследованию.
Чтобы уточнить способ размножения многобугорчатых, от которых не сохранилось никаких следов репродуктивных органов, ученые использовали метод, уже успешно зарекомендовавший себя при изучении как вымерших, так и современных видов: они сделали тончайшие срезы бедренных костей животных и исследовали их микроструктуру. Были проанализированы срезы костей 18 образцов многобугорчатых, живших на рубеже мела и палеогена (примерно 68-65,5 млн. лет назад) на территории нынешней Монтаны, США. В центре полученных срезов находилась незрелая грубоволокнистая костная ткань, в которой коллагеновые волокна идут в разных направлениях, а снаружи ее окружали слои зрелой пластинчатой костной ткани, в которой волокна расположены параллельно друг другу. Все они свидетельствуют о том, насколько быстро росло животное: незрелая кость образуется в периоды быстрого роста, а зрелая — в периоды медленного.
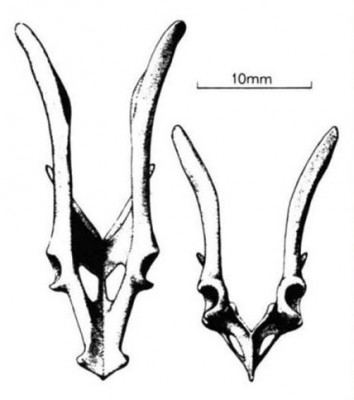
Прорисовки реконструированного таза криптобаатара (
Kryptobaatar dashzevegi): сверху (слева) и спереди (справа).
Затем ученые изучили поперечные срезы бедренных костей 35 видов мелких млекопитающих, живущих сегодня, — 28 плацентарных и 7 сумчатых. У плацентарных наблюдалась столь же выраженная слоистость, что и у многобугорчатых, а вот у сумчатых зверей картина была иной: почти вся бедренная кость целиком состояла из пластинчатой костной ткани, с узким слоем грубоволокнистой. Поскольку количество зрелой костной ткани в наружном слое кости сильно коррелирует с продолжительностью молочного вскармливания (у сумчатых оно длится намного дольше, чем у плацентарных), полученная картина явно свидетельствует, что молодые многобугорчатые развивались скорее по ''плацентарному'', чем по ''сумчатому'' сценарию. Соответственно можно предполагать, что в зародышевый период развития плацента у них была развита лучше, чем у сумчатых.
Так как внешняя зрелая костная ткань закладывается уже после рождения, по мере роста диаметра бедренной кости, можно судить о том, насколько детеныш уступает в размерах взрослому животному. У сумчатых эта разница ошеломительна — скажем, у большого рыжего кенгуру (Macropus rufus), вес которого достигает 85 кг, детеныш рождается весом всего около 0,75 г. А вот у плацентарных она выражена намного меньше: скажем, у 15-граммовой обыкновенной бурозубки (Sorex araneus) детеныш весит 0,45 г, а у 50-килограммового снежного барса (Panthera uncia) котята рождаются полукилограммовыми. Из этого можно сделать вывод, что новорожденные многобугорчатые были не настолько меньше матери, чтобы их можно было принять за зародышей, и самки сравнительно недолго кормили детенышей молоком. Сравнив толщину незрелой костной ткани у изученных видов, ученые подсчитали, что примерное время лактации многобугорчатых составляло около месяца, как и у многих нынешних грызунов.

Самка мезодмы (Mesodma sp.) с детенышами в подземном укрытии. Автор — А. Атучин.
Полученные результаты заставляют пересмотреть взгляды на репродуктивные стратегии териевых млекопитающих и бытующее мнение, что у сумчатых размножение более примитивное, а у плацентарных — прогрессивное. Скорее всего, общий предок многобугорчатых, сумчатых и плацентарных размножался ''плацентарным'' способом, который затем сохранили его потомки, либо же, что менее вероятно, и плацентарные, и многобугорчатые независимо развили такой способ размножения, при котором период молочного вскармливания (а значит, относительной маломобильности матери, вынужденной возвращаться к беспомощным детенышам) сократился за счет удлинения внутриутробного развития. По-видимому, правы были те исследователи, которые еще полвека назад предполагали, что ''сумчатый'' способ размножения является более поздней адаптацией, призванной усилить материнский контроль над репродуктивными ресурсами в непредсказуемых условиях среды. Следовательно, это не плацентарные ''улучшили'' способ размножения сумчатых, а сумчатые ''изобрели'' новую репродуктивную стратегию.
Примечательно, что некогда древнейшим представителем многобугорчатых считали Tritylodon, несмотря на крупные размеры и основываясь на поверхностном морфологическом сходстве этого цинодонта с представителями рассматриваемой группы. Самих многобугорчатых часто относили к сумчатым, а некоторые южноамериканские кайнозойские сумчатые иногда неправильно причислялись к многобугорчатым. Также высказывалось предположение, что многобугорчатые являются предками однопроходных. В пользу этой точки зрения приводились такие признаки последних, как утрата слезной и редукция скуловой кости, а также своеобразно складчатые зачаточные щечные зубы современного утконоса.
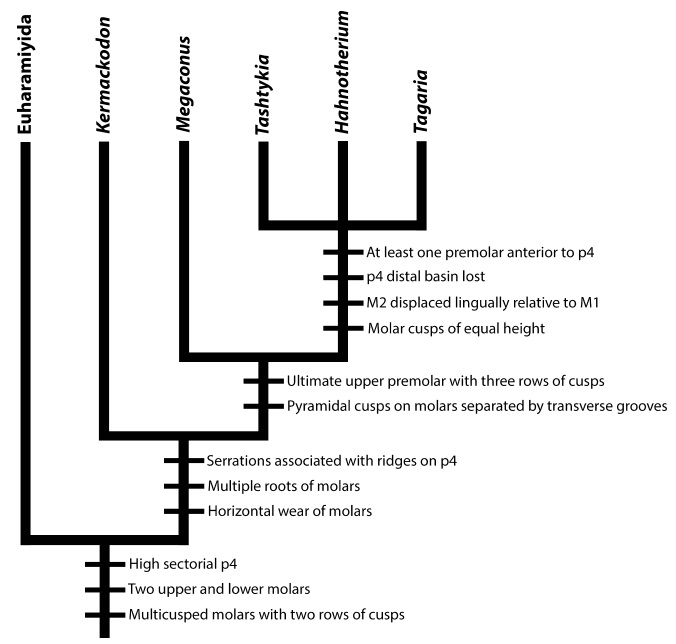
Филогенетические отношения эвхарамийид, многобугорчатых и форм с сочетанием признаков обеих групп. Отмечены особенности строения зубов, которые появляются у конкретных таксонов.
О происхождении многобугорчатых известно мало, но основная гипотеза предполагает их родство с другим отрядом аллотериев — харамийидами (Haramiyida), существовавшими с позднего триаса по поздний мел. В связи с открытым в Сибири примитивных среднеюрских многобугорчатых можно предположить, что именно здесь находился центр их происхождения. Другие древние представители отряда известны из Китая и Западной Европы. В дальнейшем многобугорчатые широко распространились по северным материкам, некогда составлявших континент Лавразию. Из числа частей суши, ранее образующих южный континент Гондвану, многобугорчатые найдены на Индостане, Мадагаскаре и в Австралии.
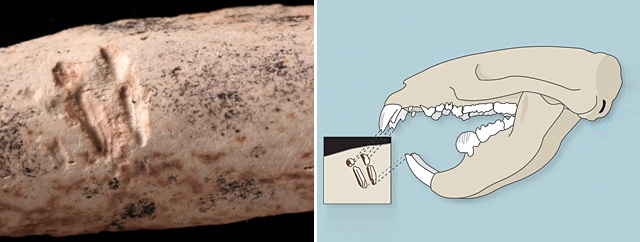
Многобугорчатые — большая, богато расчлененная группа, просуществовавшая без существенных морфологических изменений огромный промежуток времени и получившая широкое распространение. Это свидетельствует о том, что эти животные были хорошо приспособлены к окружающим условиям. Большинство представителей отряда являлись наземными, некоторые хорошо прыгали или обладали способностью к рытью, многие поздние формы вели древесный образ жизни. По-видимому, многобугорчатые тяготели к лесным стациям. Они не были способны к продолжительному быстрому бегу и для защиты от хищников использовали ядовитые шпоры на задних конечностях. Для рода Filikomys установлена жизнь небольшими группами в подземных норах.
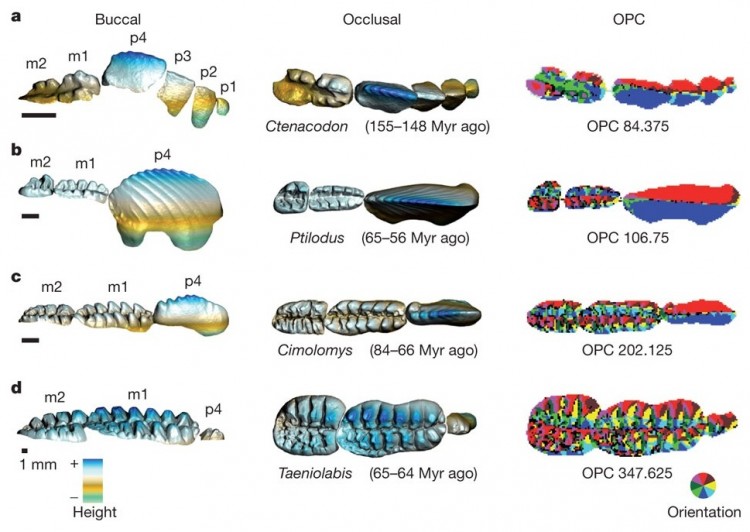
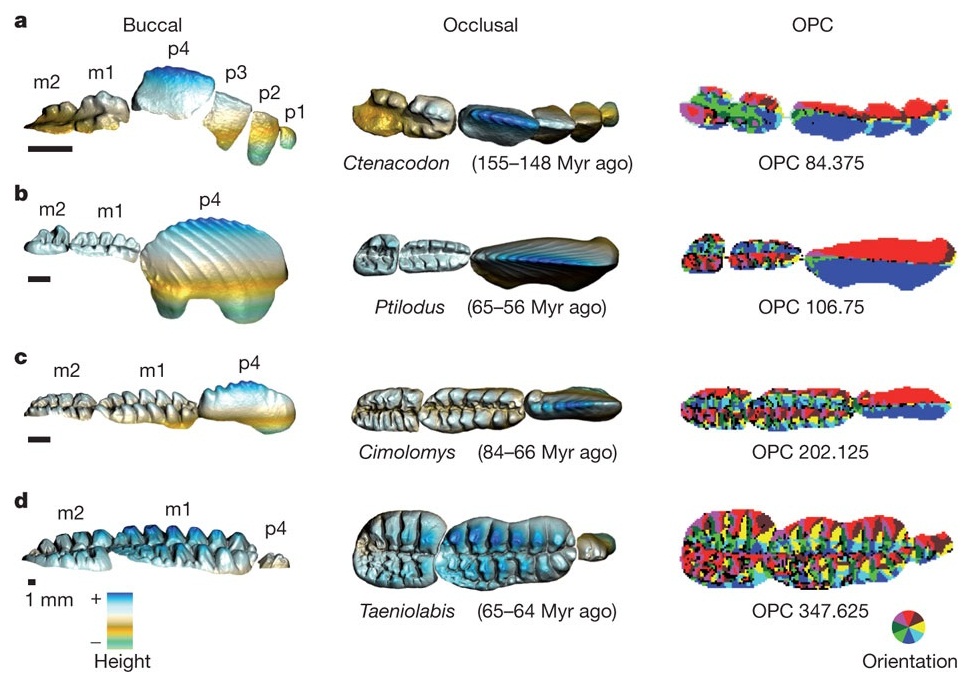
Трехмерные модели нижних щечных зубов 4 представителей многобугорчатых (сверху вниз: Ctenacodon, Ptilodus, Cimolomys, Taeniolabis). Левая колонка — вид сбоку (со стороны щеки), средняя и правая — вид сверху (жевательная поверхность). Справа разными цветами показаны площадки различной ориентации, то есть наклоненные в разные стороны. Показатель OPC (количество таких площадок) характеризует сложность зубной поверхности. m — коренные, p — предкоренные.
Пищевые предпочтения многобугорчатых долгое время оставались предметом споров: различные исследователи считали их хищными, всеядными или преимущественно растительноядными. Наибольшее признание получила последняя гипотеза. Полагали, что длинные резцы этих животных могли служить для прокалывания и прогрызания твердых плодов, большой задний нижний предкоренной — для разрезания оболочек более мягких и размельчения крупных плодов, а многобугорчатые коренные — для их раздавливания.
Затем на современных млекопитающих было продемонстрировано, что о характере их питания можно судить по показателю OPC (Orientation Patch Count), характеризующему сложность жевательной поверхности щечных зубов: чем больше на ней всевозможных бугорков и ложбинок, тем меньше доля животной и выше доля растительной пищи в рационе. Это правило достаточно строго выполняется для млекопитающих разных размеров и относящихся к разным отрядам.
Предполагаемые филогенетические связи и сравнительная морфология зубов североамериканских многобугорчатых.
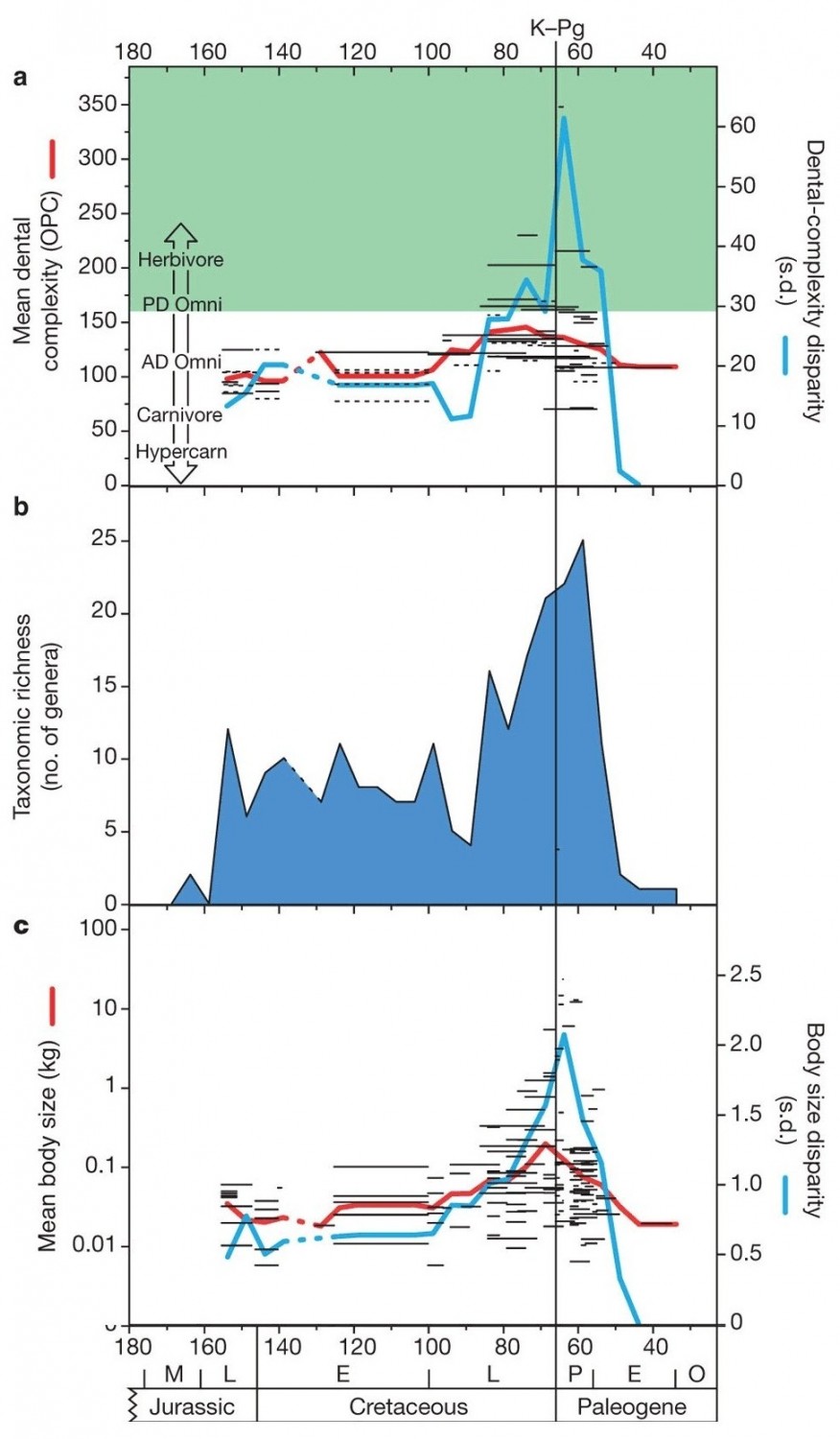
Анализ OPC многобугорчатых показал, что их юрские и раннемеловые представители, скорее всего, были всеядными животными, предпочитавшими животную пищу. Такие же значения OPC характерны для современных зверей, питающихся насекомыми и фруктами. Разброс значений оставался небольшим, что говорит об однообразии пищевых стратегий в пределах отряда многобугорчатых. Однако в позднем мелу, особенно во второй его половине, среди многобугорчатых начали появляться формы со все более сложными щечными зубами. Среднее значение OPC стало расти, что говорит о переходе многих представителей отряда на растительную диету. При этом наряду с фитофагами сохранялись и всеядные формы. Многобугорчатые стали более разнообразными по своим пищевым предпочтениям, что отражается в росте разброса (стандартного отклонения) OPC.
Во второй половине позднего мела выросло также и таксономическое разнообразие (число родов) многобугорчатых, их средний размер, а также разброс по размеру. Все эти показатели (OPC, число родов, размер) являются в значительной мере независимыми, то есть их изменения не вытекают друг из друга. Существуют статистические аргументы в пользу того, что рост среднего значения OPC в конце мела не является тривиальным следствием роста числа родов, сохранившихся в палеонтологической летописи. Хотя среди крупных многобугорчатых формы с высокими значениями OPC встречаются чаще, чем среди мелких, эта зависимость не строгая.
Погрызы на кости, оставленные многобугорчатым.
Многобугорчатые относительно легко пережили биосферный кризис на рубеже мезозоя и кайнозоя. Некоторые их семейств вымерли, но дифференциация оставшихся линий продолжалась, так что в целом родовое разнообразие в начале палеогена по сравнению с концом мела даже возросло. В палеоцене многобугорчатые оставались столь же разнообразны по размерам и питанию, как и в конце мела. В конце палеоцена и в эоцене группа пришла в упадок, ее разнообразие стало быстро сокращаться, а последние ее представители были мелкими и имели простые жевательные поверхности, подобно древнейшим представителям отряда.
Такая динамика хорошо согласуется с гипотезой о том, что многобугорчатые столкнулись с все ужесточающейся конкуренцией со стороны плацентарных — сначала кондиляртр, позднее плезиадаписообразных и, наконец, грызунов. Эти группы характеризовались более прогрессивной физиологией и поведением, чем многобугорчатые. Освоив те же экологические ниши, они научились использовать их ресурсы более эффективно. Дополнительный прессинг на многобугорчатых могли создавать климатические изменения и появление новых хищников.
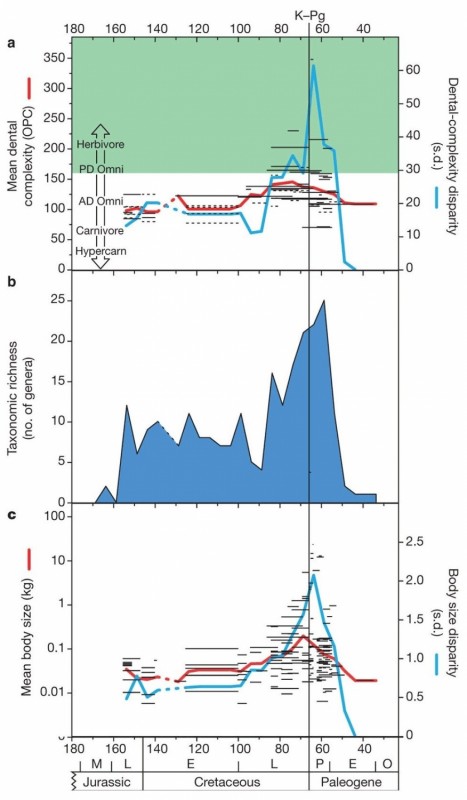
Изменения морфологического и таксономического разнообразия многобугорчатых во времени. По горизонтальной оси — время в млн лет назад. Вертикальной черной линией показан рубеж мела и палеогена (65,5 млн. лет назад), когда произошло массовое вымирание. a — сложность зубной поверхности (OPC). Черными горизонтальными линиями показаны данные по отдельным родам, красная линия — среднее значение OPC, синяя — разброс (стандартное отклонение), характеризующий разнообразие типов питания. Зеленым закрашена область значений OPC, характерная для зверей, предпочитающих растительную пищу (Herbivore — растительноядные, PD Omni — всеядные с преобладанием растительной пищи). Незакрашенная область соответствует преобладанию животной пищи в рационе (от AD omni — всеядных с уклоном в хищничество до Hypercarn — "гиперхищников"). b — число родов. c — масса тела (кг).
Судя по всему, главным стимулом для быстрого роста разнообразия многобугорчатых и сдвига их пищевых предпочтений в сторону фитофагии была экспансия цветковых растений. Она началась несколько раньше, в середине мела, но именно во второй половине позднего мела происходил особенно быстрый рост их разнообразия и численности. Среди цветковых в то время появилось множество быстрорастущих травянистых форм, потенциально представляющих собой ценнейший пищевой ресурс. Многобугорчатые сумели его освоить, что и привело к росту их разнообразия и численности. Поскольку цветковые растения мало пострадали во время кризиса, кормовая база многобугорчатых не была подорвана, и это помогло им сохранить высокое разнообразие в палеоцене. Дополнительными факторами, обеспечившими выживание группы, могли быть теплокровность, небольшой размер и, соответственно, высокая численность популяций, а также достаточно эффективная забота о потомстве. В общей сложности многобугорчатые просуществовали около 120 млн. лет — своеобразный рекорд среди отрядов млекопитающеобразных. Это первая группа тетрапод, достигшая доминирования в адаптивной зоне "грызунов" — т. е. небольших фитофагов.
По мнению ряда исследователей, расцвет многобугорчатых мог быть одной из косвенных причин упадка динозавров. Многобугорчатые были фактически первыми высокоэффективными мелкими фитофагами среди наземных позвоночных. Поскольку растительной пищи в общем случае во много раз больше, чем животной, популяции мелких фитофагов могут достигать очень высокой численности (яркий пример — современные грызуны). Появление множества мелких растительноядных млекопитающеобразных, в свою очередь, создало предпосылки для эволюции специализированных охотников на этих животных, то есть хищников, специализирующихся на мелкой четвероногой добыче. Однажды появившись, такие хищники, несомненно, стали бы представлять смертельную угрозу для маленьких детенышей динозавров, которых гигантские родители едва ли могли эффективно охранять.
Состав отряда следующий:
семейство †Ханотериевые (Hahnotheriidae Butler et Hooker, 2005);
семейство †Кермакодонтовые (Kermackodontidae Butler et Hooker, 2005);
подотряд †Плагиаулациды (Plagiaulacida Simpson, 1925);
надсемейство †Аллодонтоиды (Allodontoidea Marsh, 1889);
семейство †Аллодонтовые (Allodontidae Marsh, 1889);
семейство †Софиябаатаровые (Zofiabaataridae Bakker, 1992);
надсемейство †Паульхоффатиоиды (Paulchoffatioidea Hahn, 1969 sensu Hahn et Hahn, 2003);
семейство †Паульхоффатиевые (Paulchoffatiidae Hahn, 1969);
семейство †Пинейродонтовые (Pinheirodontidae Hahn et Hahn, 1999);
надсемейство †Плагиаулацоиды (Plagiaulacoidea Ameghino, 1894);
семейство †Плагиаулациевые (Plagiaulacidae Gill, 1872);
семейство †Эобаатаровые (Eobaataridae Kielan-Jaworowska, Dashzeveg et Trofimov, 1987);
семейство †Альбионбаатаровые (Albionbaataridae Kielan-Jaworowska et Ensom, 1994);
подотряд †Цимолодонты (Cimolodonta McKenna, 1975);
семейство †Коррьебаатаровые (Corriebaataridae Rich, Vickers-Rich, Flannery, Kear, Cantrill, Komarower, Kool, Pickering, Trusler, Morton, Van Klaveren et Fitzgerald, 2009);
семейство †Боффиевые (Boffidae Hahn et Hahn, 1983);
семейство †Цимоломовые (Cimolomyidae Marsh, 1889);
семейство †Когайоновые (Kogaionidae Radulescu et Samson, 1996);
семейство †Эвкосмодонтовые (Eucosmodontidae Jepsen, 1940);
семейство †Микрокосмодонтовые (Microcosmodontidae Holtzman et Wolberg, 1977);
надсемейство †Птилодонтоиды (Ptilodontoidea Cope, 1887);
семейство †Цимолодонтовые (Cimolodontidae Marsh, 1889);
семейство †Неоплагиаулациевые (Neoplagiaulacidae Ameghino, 1890);
семейство †Птилодонтовые (Ptilodontidae Cope, 1887);
надсемейство †Джадохтатериоиды (Djadochtatherioidea Kielan-Jaworowska et Hurum, 1997);
семейство †Хулсанбаатаровые (Chulsanbaataridae Kielan-Jaworowska, 1974);
семейство †Слоанбаатаровые (Sloanbaataridae Kielan-Jaworowska, 1974);
семейство †Джадохтатериевые (Djadochtatheriidae Kielan-Jaworowska et Hurum, 1997);
надсемейство †Тениолабидоиды (Taeniolabidoidea Granger et Simpson, 1929);
семейство †Тениолабидовые (Taeniolabididae Granger et Simpson, 1929);
семейство †Лямбдопсаловые (Lambdopsalidae Chow et Qi, 1978).
Ссылки
Древнейшие многобугорчатые млекопитающие юрского периода и Сибирь как центр происхождения мультитуберкулят (PALEO.RU)
перейти Марков А. Экспансия растительноядных млекопитающих началась задолго до вымирания динозавров. — 2006 (Элементы.Ру)
перейти Многобугорчатые (Википедия)
перейти Multituberculata (Paleobiology Database)
перейти
Литература
Зоогеография палеогена Азии / Отв. ред. Крамаренко Н. Н. — М.: Наука, 1974
Иванов А. О., Черепанов Г. О. Ископаемые высшие позвоночные. — СПб.: Издательство Санкт-Петербургского университета, 2001
Кэрролл Р. Палеонтология и эволюция позвоночных: В 3-х т. Т. 2. — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 13: Млекопитающие / Гл. ред. Орлов Ю. А. — М.: Государственное научно-техническое издательство литературы по геологии и охране недр, 1962
Павлова М. В. Палеозоология. Ч. 2. Позвоночные. — М.-Л.: Государственное издательство, 1929
Ромер А. Ш. Палеонтология позвоночных. — М. — Л.: Государственное научно-техническое издательство нефтяной и горно-топливной литературы, 1939
Россолимо О. Л., Павлинов И. Я., Крускоп С. В., Лисовский А. А., Спасская Н. Н., Борисенко А. В., Панютина А. А. Разнообразие млекопитающих. Ч. 1-3. — М.: Изд-во КМК, 2004