Морганукодоны — Morganucodon
Клада Млекопитающеобразные (Mammaliaformes)
Отряд †Морганукодонты (Morganucodonta)
Семейство †Морганукодонтовые (Morganucodontidae)
Род †Морганукодоны (Morganucodon)
Вид †Morganucodon peyeri
Вид †Morganucodon watsoni
Вид †Morganucodon tardus
Вид †Morganucodon oehleri
? Вид †Morganucodon heikuopengensis
Морганукодоны (Morganucodon Kühne, 1949) — род семейства морганукодонтовых (Morganucodontidae Kühne, 1958), происходящий из позднего триаса - средней юры Европы, позднего триаса - ранней юры Азии и ранней юры Северной Америки. Установлены 4 или 5 видов.

Реконструкция внешнего облика морганукодона (Morganucodon watsoni). Автор — FunkMonk.
Синонимы: Eozostrodon Parrington, 1941 (частью); Rostrodon Young, 1978.
Morganucodon был описан В.Г. Кюне в 1949 г. по фрагментарным остаткам, в изобилии встречающимся в карстовых расселиннах графства Гламорган на территории Уэльса в Великобритании. Родовое наименование в переводе означает «зуб из Гламоргана».
В отличие от большинства других базальных млекопитающеобразных, ископаемый материал Morganucodon весьма многочисленный. Из осадка, заполняющего карстовые трещины в Великобритании, происходят тысячи разрозненных элементов, а в Китае помимо отдельных костей найден сочлененный материал. Это позволило восстановить и детально изучить череп и большую часть посткраниального скелета.
Размеры мелкие: длина черепа составляет 2-3 см, общая длина оценивается в 10-15 см, вес — приблизительно в 20 г.
Череп

Реконструированный череп с нижней челюстью Morganucodon.
Череп длиной относительно тела кажется крупным. По сравнению с черепами цинодонтов Thrinaxodon и Probainognathus он длиннее и тоньше. Скуловая дуга узкая, выгнута дорсально, но меньше выдается наружу. Височное окно и глазница слились. Мозговая коробка выглядит гораздо крупнее, чем у любого из известных цинодонтов, что отчасти можно объяснить более мелкими размерами всего черепа (менее половины его длины у Probainognathus).
Теменные кости слившиеся; линия их слияния образует узкий сагиттальный гребень. С утратой заглазничных и предлобных костей лобные становятся гораздо более крупными, чем у цинодонтов, элементами и распространяются вентролатерально по дорсальной части мозговой коробки, как и теменные кости. Спереди от глазниц они смыкаются с дорсальными отростками небных костей. В отличие от териевых (сумчатых и плацентарных) млекопитающих чешуйчатые кости в образовании мозговой коробки не участвуют.
Хотя конец морды известен плохо, можно полагать, что наружные ноздри сливались в одно отверстие. Вентрально предчелюстные и верхнечелюстные кости обрамляют резцовое отверстие, расположенное чуть латеральнее средней линии. У современных млекопитающих оно связано с якобсоновым органом, добавочным выростом носовой полости, служащим для восприятия запаха содержимого рта. Над вторичным небом находится обширная носовая полость. По внутренним поверхностям верхнечелюстных и носовых костей проходят продольные гребни, к которым при жизни прикреплялись турбиналии, выстланные обонятельным эпителием.
Зубной ряд распространяется, как и вторичное небо, более чем на половину длины черепа. Расстояние между задним концом последнего и вторичным небом по сравнению с цинодонтами уменьшено и у настоящих млекопитающих сокращается еще сильнее. Размер крыловидных костей уменьшается, но на них еще сохраняется латеральный выступ, ограничивающий медиальные движения нижней челюсти. Возможно, у Morganucodon еще была наружная крыловидная кость (ectopterygoideum), утраченная большинством млекопитающих. Квадратная ветвь крыловидной кости идет назад в виде узкого отростка снаружи от надкрыловидной полости, открытой вентрально латеральнее основной клиновидной кости.
Наружная поверхность задней части мозговой коробки в целом напоминает таковую у хищных цинодонтов, но околоушная, или каменистая, кость заходит вперед относительно дальше, чем у Probainognathus, и окружает отверстия для максиллярной и мандибулярной ветвей тройничного нерва, которые проходят у цинодонтов между околоушной и крылоклиновидной костями, а у современных териевых млекопитающих — сквозь крылоклиновидную кость. У сумчатых и плацентарных эти отверстия называются соответственно круглым (foramen rotundum) и овальным (f. ovale). Поскольку у Morganucodon они находятся не на крылоклиновидной, а на каменистой кости, то называются ложнокруглым (f. pseudorotundum) и ложноовальным (f. pseudoovale).
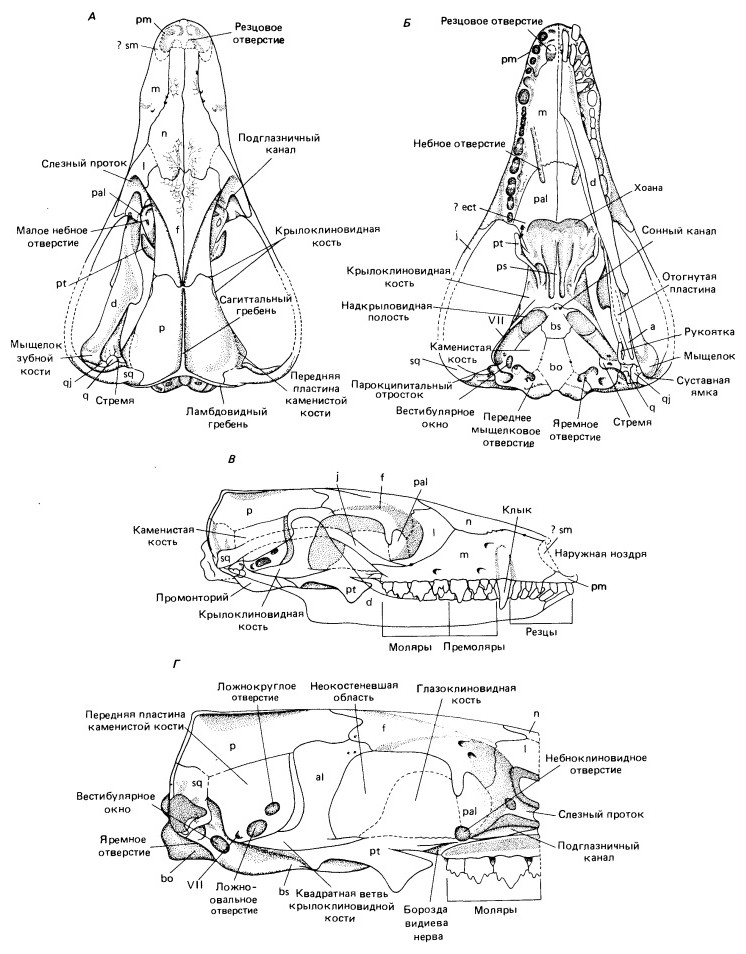
Прорисовки частично реконструированного черепа с нижней челюстью Morganucodon: А — сверху; Б — со стороны неба; В — сбоку; Г — латеральный вид мозговой коробки с удаленной скуловой дугой. Длина оригинала 2 см. Крылоклиновидная кость у образца сломана, поэтому представляется неестественно узкой.
Крылоклиновидная кость распространяется назад в виде узкой квадратной ветви под каменистой костью примерно до заднего конца ее передней пластинки. Крупное отверстие в боковой стенке черепа медиальнее глазницы, сохранявшееся у хищных цинодонтов, отчасти закрывается новой, глазоклиновидной (orbitosphenoideum), костью, через которую проходит зрительный (II-й) нерв. У некоторых Tritylodontidae боковая стенка окостеневает более полно, так что по этому признаку они продвинутее базальных млекопитающеобразных.
У мозговой коробки широкое треугольное основание. Основная клиновидная кость прочно связана швом с крыловидной, но с дорсальной стороны можно видеть базиптеригоидный отросток, ранее образовывавший подвижное сочленение с крыловидной костью; здесь основная клиновидная кость обособлена от остального черепа. Спереди она сливается с узким отростком парасфеноида (у большинства млекопитающих парасфеноид остается самостоятельным). Спереди он продолжается тонким срединным сошником.
Вентрально в основной клиновидной кости расположены сонные отверстия (f. caroticum) для прохождения сонных артерий. Ход их у млекопитающих довольно разнообразен. У Cynognathus, Gomphodontia и Tritylodontidae сонные артерии входят в турецкое седло с боков.
Слуховая капсула у Morganucodon, как и у всех млекопитающеобразных, окостеневает единым образованием — околоушной, или каменистой, костью, возникающей при слиянии заднеушной и переднеушной костей более примитивных тетрапод. Вентрально она формирует широкое дно мозговой коробки, тесно связанное с основной клиновидной и основной затылочной костями.
Важное отличие от цинодонтов — разрастание вперед основания каменистой кости с образованием части дна надкрыловидной полости медиальнее вертикально ориентированной передней пластины каменистой кости (передний отросток околоушной кости в менее развитом виде свойствен всем цинодонтам). С дорсальной стороны можно видеть крупное углубление для полулуного ганглия V-го нерва. Сзади и дорсальнее находится второе заметное углубление на внутренней поверхности каменистой кости — поддуговая яма (fossa subarcuata), где располагается боковой придаток мозжечка — часть мозга, играющая важную роль в координации мышечных сокращений.
Улитка внутреннего уха гораздо крупнее, чем у цинодонтов, и ее положение можно определить снаружи по заметному выступу — промонторию (promontorium). Как у терпсид и однопроходных, она не завита спирально в отличие от сумчатых и плацентарных. Вентрально слуховая капсула прободена кохлеарным (улиточным) отверстием (f. cochlearis) и вестибулярным (преддверным) отверстием (f. vestibularis), или овальным окном (f. ovalis), на которое опирается основание стремени.
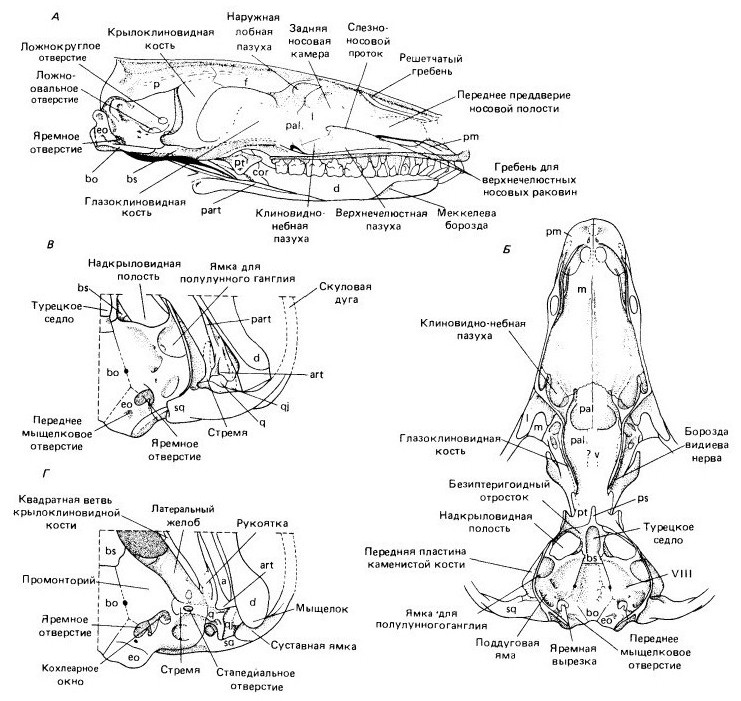
Прорисовки черепного материала Morganucodon: А — сагиттальный разрез черепа, медиально; Б — дно черепа, сверху; В — задняя часть мозговой коробки и области челюстного сустава, сверху; Г — область среднего уха и челюстного сустава, снизу.
Эта область мозговой коробки значительно отличается от свойственной продвинутым плацентарным млекопитающим, поскольку здесь еще нет слуховой буллы, которая может сформироваться только после исключения сочленовной и квадратной костей из челюстного сустава. У примитивных сумчатых слуховая булла также отсутствует, т. е. строение остается примитивным.
Дорсальный край теменной и чешуйчатой костей формирует поперечный ламбдовидный гребень, отделяющий затылочную поверхность от височного окна. В отличие от продвинутых млекопитающих шовные соединения костей затылка еще хорошо заметны. Мыщелки полностью образованы боковыми затылочными костями, а основная затылочная видна сзади только узкой полоской. По сравнению с современными териевыми млекопитающими мыщелки располагаются довольно низко. Непосредственно спереди от каждого из них находится крошечное переднее мыщелковое отверстие для XII-го черепномозгового нерва, а между боковой затылочной и каменистой костями — отверстие для внутренней яремной вены, языкоглоточного (IX-го), блуждающего (X-го) и добавочного (XI-го) нервов, называемое у рептилий яремным, а у млекопитающих — задним рваным отверстием (f. lacerum posterior).
Между чешуйчатой и каменистой костями расположена задневисочная яма, утраченная у более поздних млекопитающих. Таблитчатых и верхнезатылочных костей нет ни у одного из изученных экземпляров, однако полагают, что они оставались самостоятельными центрами окостенения, как и заднетеменные кости на ранних стадиях развития современных млекопитающих.
Челюстной сустав базальных млекопитающеобразных располагается по уровню непосредственно перед затылочным мыщелком, но у продвинутых форм смещен вперед. Его примитивное заднее положение, возможно, объясняется унаследованной от цинодонтов тесной связью между постдентальными нижнечелюстными костями и слуховой капсулой. У Morganucodon чешуйчатая кость образует отчетливую, но неглубокую суставную впадину для сочленения с мыщелком зубной кости. Непосредственно медиальнее ее находится углубление для квадратноскуловой и квадратной костей, образующих сустав с сочленовной. Квадратная кость медиально поддерживается парокципитальным отростком слуховой капсулы. Челюстной сустав располагается немного выше уровня зубного ряда, тогда как у хищных цинодонтов — вентральнее его.
Стремя пронизано крупным стапедиальным отверстием и сочленяется с медиальной поверхностью квадратной кости. Эти элементы, по-видимому, уже функционировали как слуховые косточки, но по своим относительным размерам они лишь немного мельче, чем у цинодонтов. Судя по величине угловой кости и ретроартикулярного отростка сочленовной, барабанная перепонка должна была оставаться крайне крупной по сравнению с таковой у современных млекопитающих. Показано, что соотношение размеров барабанной перепонки и подошвы стремени могло обеспечивать их функционирование в качестве эффективной структуры выравнивания импедансов, но из-за их величины и сохраняющейся тесной связи с нижнечелюстным аппаратом эти элементы, вероятно, передавали звук гораздо хуже, чем у современных млекопитающих.
Нижняя челюсть
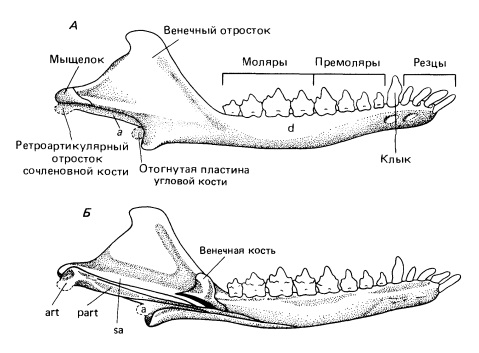
Прорисовки частично реконструированной нижней челюсти Morganucodon: А — снаружи; Б — изнутри.
Нижняя челюсть Morganucodon снаружи выглядит типично для млекопитающих: крупная зубная кость образует высокий венечный отросток и выраженный мыщелковый отросток для сочленения с черепом. Как и у продвинутых цинодонтов, ее нижняя граница оканчивается сзади округлым отростком, который сравнивали с угловым у териевых млекопитающих, несмотря на то что он находится спереди от мыщелка. Такое же его расположение, свойственное и другим ранним млекопитающеобразным, особенно Dinnetherium, предполагает, что данная структура примитивнее и поэтому лучше называть ее ложноугловым отростком (processus pseudoangularis). Между ним и мыщелком заметны узкая отогнутая пластинка угловой кости и рукоятка сочленовной, которые должны были поддерживать обширную барабанную перепонку.
С медиальной стороны видно, что все постдентальные кости цинодонтов еще сохранены, хотя их размер сильно редуцирован. Стержневидные предсочленовная, угловая и надугловая кости лежат в углублении на внутренней поверхности зубной. Венечная кость представляет собой плоскую, асимметричную пластинку у основания венечного отростка. Суставная поверхность сочленовной кости вогнута и направлена почти прямо назад, чтобы окружать обращенное вперед основание квадратной кости. Симфиз нижней челюсти развит несильно, и обе ветви могут двигаться независимо друг от друга.
Зубы
Зубная формула довольно изменчива: I3-5/i4-5, Cl/c1, Р4-5/p4-5, М3-4/m3-5. Подобно типичным млекопитающим, резцы, клыки и предкоренные зубы сменялись только один раз в жизни, а коренные — никогда. Коренные зубы с 2 корнями и 3 основными расположенными линейно бугорками, окклюзирующие.
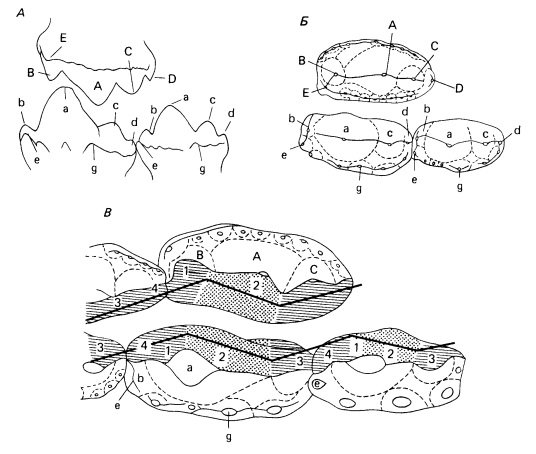
Прорисовки зубного материала Morganucodon. А, Б — верхний и нижние коренные изнутри и с жевательной стороны. В — 2 верхних и 3 нижних коренных с жевательной стороны. Стирающиеся грани образуют две стороны «широкоугольных», противоположно направленных треугольников. Ориентация этих граней показана жирными черными линиями. Передняя сторона — слева.
Характер износа зубов Morganucodon очень специфичен и тесно коррелирует с их строением. Как у Galesauridae и у Chiniquodontidae, коронки латерально сжаты и спереди назад несут ряд апикальных бугров. Расположение и функции главных из них постоянны. Бугры верхних зубов условно обозначаются заглавными буквами, а нижних — строчными. Самый высокий бугор приблизительно посередине зуба будет соответственно — А, а. Бугор перед ним — В, b. Бугры С, с и D, d расположены сзади А, а. У Galesauridae и некоторых Chiniquodontidae вдоль внутреннего, или лингвального, края коренных проходит низкий гребень, цингулюм (поясок). У Morganucodon помимо этого есть еще латеральный, или буккальный, цингулюм на верхних зубах. На нем расположены мелкие бугорки.
Бугры острые и могли использоваться для прокусывания добычи с момента прорезания зубов, но на только что прорезавшихся зубах участки, пригодные для этого, ограничены. На протяжении жизни животного в результате специфического характера стирания между верхними и нижними зубами развивались более эффективные режущие поверхности. Медиальная поверхность А (главный бугор верхнего моляра) снашивается о латеральную поверхность нижнего зуба между а и с; латеральная поверхность а — о медиальную между А и В. В дальнейшем в процессе истирания устанавливается зигзагообразное расположение фасеток снашивания, максимально эффективное при скусывании. Этот характер строения наиболее четко виден с окклюзионной стороны. Геометрия фасеток снашивания показывает, что зубы нижней челюсти должны были двигаться при смыкании и медиально, и дорсально.
По таким примитивным признакам, как линейное расположение 3 главных бугров коренных, присутствие на этих зубах лингвального цингулида и связанных с ним выступов, а также наличие лингвальных и буккальных бугорков на зубной поверхности Morganucodon отличается от Eutriconodonta. Соседние нижние коренные совмещаются посредством расположения дистального острого выступа в выемке между выступами последующего зуба, отличая таким образом Morganucodon от Dinnetherium, Kuehneotherium и Eutriconodonta. Для Morganucodon характерен специфический тип окклюзии коренных зубов: центральный бугор нижнего коренного заходит с лингвальной стороны во впадину между буграми верхнего. Центральный бугор верхнего коренного на буккальной стороне заходит во впадину между острыми буграми нижнего. Особенностями окклюзии Morganucodon отчетливо отличается от Megazostrodon, Brachyzostrodon и Indozostrodon.
Посткраниальный скелет
По-видимому, наиболее велики отличия Morganucodon от всех терапсид в строении позвоночника. Как и у позднейших млекопитающих, шейные позвонки характеризуются очень широким невральным каналом, что, возможно, связано с большим, чем у терапсид, развитием плечевого нервного сплетения. В отличие от более продвинутых форм шейные ребра у Morganucodon не сливаются с позвонками. Впоследствии приросшие к ним основания шейных ребер окружают поперечное отверстие для шейной артерии. Дуги атланта остаются парными и не слиты с гипоцентром, но проатлант и зигапофизные сочленения с эпистрофеем утрачены, в результате чего возможно свободное вращение, как у современных млекопитающих. Тела позвонков по существу платицельные, по сохраняют следы хордальных впадин.
Грудные и поясничные позвонки отчетливо различаются. Последние не несут ребер, как и у современных млекопитающих, и их зигапофизы наклонены под углом 35-45°, что предотвращает скручивание, в отличие от почти плоских зигапофизов в грудном отделе. Остистые отростки низкие, но в их строении заметна типичная для млекопитающих особенность: на передних позвонках они наклонены назад, а на более задних — вперед. В задней части грудного отдела находятся 3 переходных, или антиклинальных, позвонка. Такое строение туловищного отдела позвоночника у млекопитающих связано с развитием сгибания — растягивания в сагиттальной плоскости в отличие от латерального изгибания у более примитивных амниот. Точное число предкрестцовых позвонков установить невозможно, но, вероятно, их было около 27; крестцовых — 2 или, возможно, 3; хвостовых — приблизительно 12. Хвост достаточно длинный и тонкий.
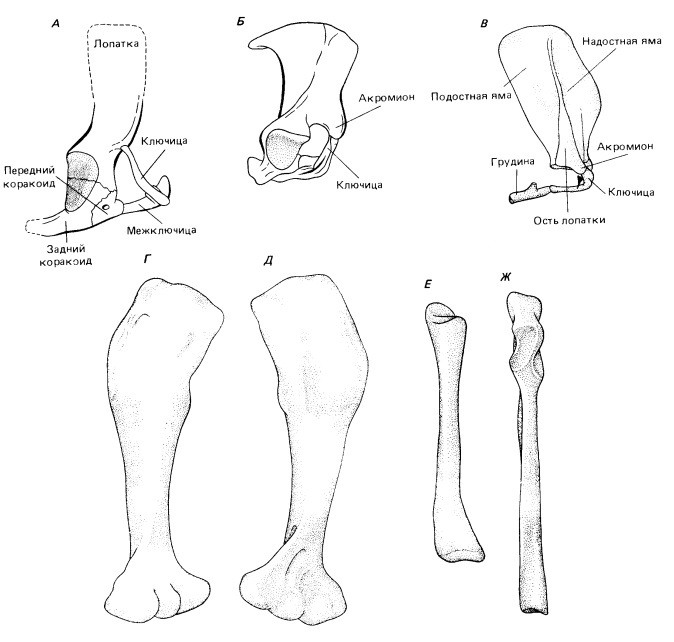
Прорисовки плечевого пояса и элементов передней конечности примитивных млекопитающих. Плечевой пояс: А — Morganucodon, Б — Tachyglossus; В — Didelphis (иллюстрирующий типичное для териев состояние с мощной остью, отделяющей переднюю надостную яму от задней подостной). Левая плечевая кость Morganucodon: Г — сверху; Д — снизу. Е, Ж — правая лучевая и левая локтевая кости того же рода, спереди.
Конечности и их пояса напоминают таковые у продвинутых цинодонтов (например, у Oligokyphus), но ряд их признаков характерен исключительно для млекопитающих. В плечевом поясе прокоракоид еще сохраняется, хотя и совершенно не участвует в образовании гленоидной впадины. Как и у цинодонтов, и однопроходных, задний коракоид несет мощный, направленный назад отросток. Лопатка, как и у продвинутых цинодонтов, гораздо примитивнее, чем у современных сумчатых и плацентарных. У териевых млекопитающих это широкая костная пластина, разделенная вертикально проходящей остью на переднюю надостную (fossa supraspinus) и заднюю подостную (f. infraspinus) ямы. Эти поверхности заняты надостной (m. supraspinatus) и подостной (m. infraspinatus) мышцами, стабилизирующими плечевой сустав у современных млекопитающих, у которых гленоидная впадина широко раскрыта. У продвинутых цинодонтов, триасовых млекопитающеобразных и однопроходных надостная мышца крепится на антеромедиальной поверхности лопаточной пластины, передний край которой гомологичен ости лопатки териевых форм. Область, лежащая у сумчатых и плацентарных под этим мускулом, — новая область окостенения спереди от исходного края лопатки. Несмотря на это, ключица продолжает сочленяться с акромионом в прежнем положении.
В реконструкции подразумевается присутствие грудины. Грудинные элементы описаны у Tritylodontidae и, предположительно, были у всех ранних млекопитающеобразных. Ключица и межключица у них еще остаются крупными; межключица сохранилась у современных однопроходных, но утрачена териевыми формами.
Как это ни странно, у Morganucodon нет признаков отдельных эпифизных окостенений. Они характерны для всех современных млекопитающих и должны были бы обнаруживаться у ранних форм, которым уже свойственны хорошо сформированные суставные поверхности и, судя по малым размерам, строго регулируемый рост.
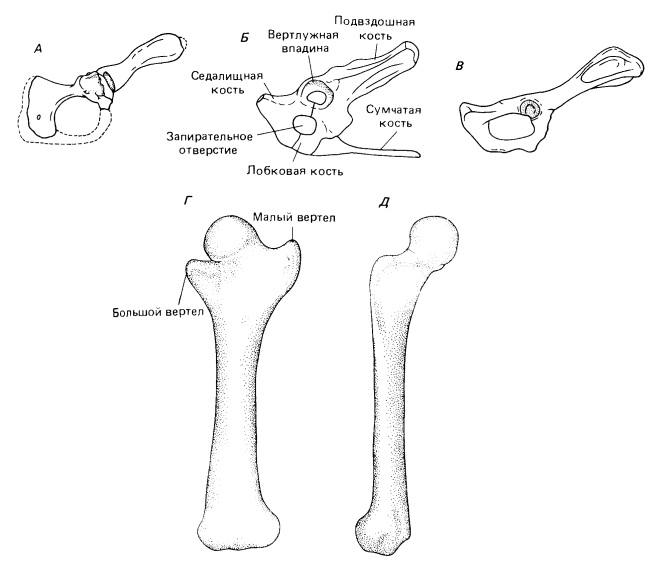
Тазовый пояс и бедренная кость примитивных млекопитающих. Таз: А — Morganucodon; Б — Tachyglossus; В — Tupaia. Левая бедренная кость Morganucodon: Г — сверху; Д — изнутри.
Конечности по отношению к туловищу выглядят тонкими. Плечевая кость остается в целом примитивной, с широко разросшимися концами, повернутыми приблизительно под углом 50 ° друг к другу. Эктэпикондилярное отверстие цинодонтов утрачено, но крупное энтэпикондилярное сохраняется. Полушаровидная головка сильно отклонена на конце кости дорсально.
В тазу запирательное отверстие крупнее, а лобковая кость мельче, чем у цинодонтов. Вертлужная впадина сохраняет рудименты отдельных сочленовых поверхностей, сформированных лобковой, подвздошной и седалищной костями, которые у более поздних млекопитающих сливаются. Нижний край вертлужной впадины неполный. Предлобковые (сумчатые) кости не найдены, но поскольку они встречаются и у Tritylodontidae, и у нескольких групп примитивных млекопитающих, то, возможно, были и у Morganucodon.
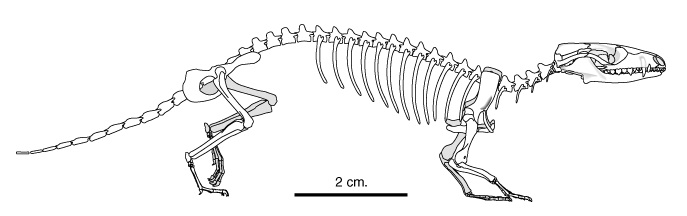
Прорисовка реконструированного скелета Morganucodon.
Бедренная кость гораздо ближе к типу строения, характерному для млекопитающих, чем плечевая. Почти сферическая головка вполне четко отделена от длинного и узкого диафиза. Углубление вблизи центра головки свидетельствует о присутствии связки (ligamentum capitis femoris), которая у современных млекопитающих служит для прикрепления кости к вертлужной впадине. Большой и малый вертелы отходят медиально и латерально от проксимального конца диафиза в той же плоскости, что и дистальная сочленовная поверхность.
Конфигурация суставных поверхностей нижних частей конечностей указывает, что постановка и характер движения последних напоминали свойственные примитивным ныне живущим небегающим млекопитающим типа сумчатого Didelphis и плацентарного Tupaia. Голеностопный сустав мало продвинут по сравнению с цинодонтным, таранная кость в отличие от современных млекопитающих находится еще не вполне дорсально по отношению к пяточной. Конечности опираются на всю стопу.

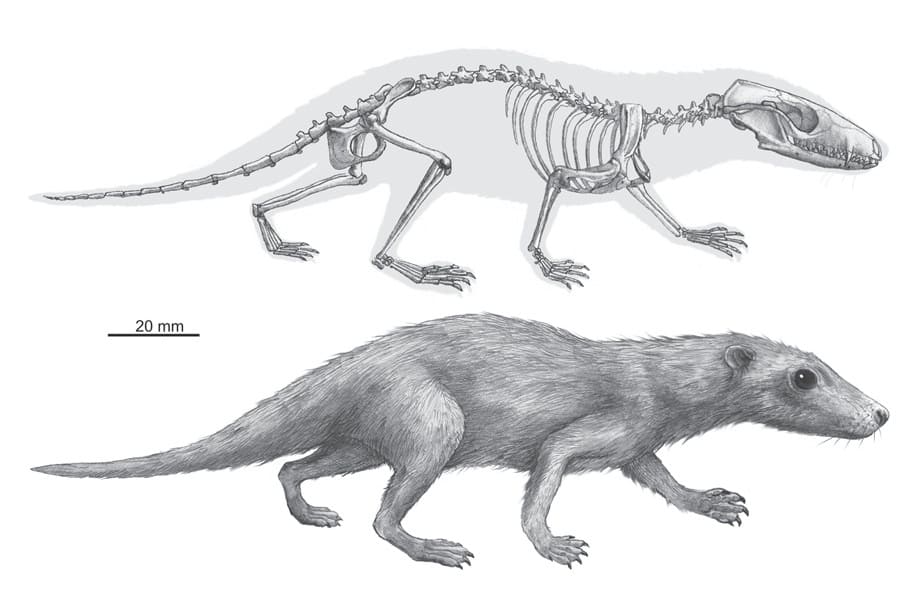
Прорисовка реконструированного скелета, вписанная в телесные очертания, и реконструкция внешнего облика морганукодона (Morganucodon sp. B). Автор — А. Hołda-Michalska.
Головной мозг
Коэффициент энцефализации Morganucodon составляет 0,32 против 0,16-0,23 у таких базальных цинодонтов, как Thrinaxodon и Diademodon. Сильнее всего увеличились обонятельная луковица и обонятельная кора головного мозга. Это со всей очевидностью указывает на развитое обоняние. Полушария переднего мозга стали выпуклыми благодаря развитию неокортекса; они закрывают средний мозг и эпифиз при взгляде сверху, как у млекопитающих. Передний мозг Morganucodon шире мозжечка, хотя мозжечок тоже заметно вырос по сравнению с базальными цинодонтами. Увеличение мозжечка говорит об улучшенной координации движений. На это же указывает и более толстый, чем у базальных цинодонтов, спинной мозг. Изменения в строении внутреннего уха свидетельствуют также о возможном улучшении слуха.
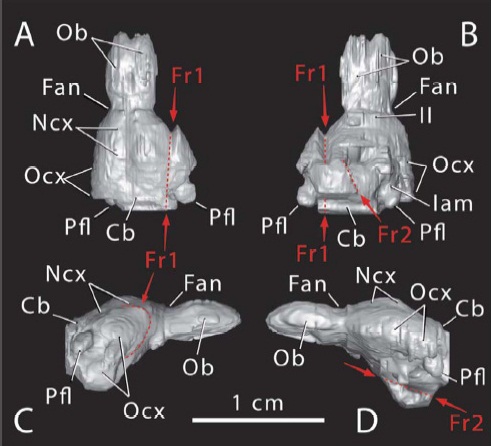
Эндокаст морганукодона (Morganucodon oehleri), реконструированный при помощи рентгеновской томографии. Ob — обонятельные луковицы, Ncx — неокортекс, Ocx — обонятельная кора, Cb — мозжечок.
Состав
К роду относят 4-5 видов.
Morganucodon peyeri Clemens, 1980 — вид, известный из позднего триаса (рэтский ярус) Швейцарии. Найден фрагментарный челюстной материал и изолированные зубы.
Morganucodon watsoni Kühne, 1949 (тип) — типовой вид, известный из ранней юры (геттангский / синемюрский ярус) Великобритании (Уэльс). Синоним: Eozostrodon watsoni (Kühne, 1949). Найдены многочисленные изолированные зубы, челюстные фрагменты с зубами и фрагменты костей посткраниального скелета. Зубная формула: I3/i4, Cl/c1, Рm4/pm4, М4/m5.

Фрагмент нижней челюсти и щечной зуб морганукодона (Morganucodon watsoni).
Morganucodon tardus Butler and Sigogneau-Russell, 2016 — вид, известный из средней юры (батский ярус) Великобритании (Юго-Западная Англия).
Morganucodon oehleri Rigney, 1963 — вид, известный из ранней юры (нижние слои формации Люфенг, геттангский - синемюрский ярусы) Китая (провинция Юньнань). Синонимы: ? Morganucodon heikuopengensis Young, 1978; Eozostrodon oehleri (Rigney, 1963) Jenkins et Crompton, 1979. Найдены практически полные черепа и некоторые элементы посткраниального скелета. Зубная формула: I4/i4, Cl/c1, Рm4/pm5, М3/m3-4.
? Morganucodon heikuopengensis Young, 1978 — сомнительный вид, известный из ранней юры (нижние слои формации Люфенг, синемюрский ярус) Китая (провинция Юньнань). Синонимы: Eozostrodon heikuopengensis (Young, 1978); Rostrodon heikuopengensis Young, 1978. Возможно, данный таксон сам является младшим синонимом Morganucodon oehleri. Найден фрагментарный челюстной материал и изолированные зубы. Зубная формула: I5/i5, Cl/c1, Рm4-5/pm4-5, М3/m3.
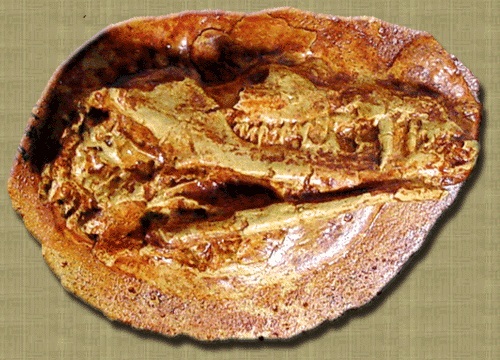
Слепок неполного черепа с нижней челюстью морганукодона (Morganucodon sp.) из ранней юры США.
Остатки, определенные лишь до рода и обозначенные Morganucodon spр., известны из позднего триаса (рэтский ярус) Швейцарии и Франции (регион Гранд-Эст), ранней юры (геттангский / синемюрский ярус) Великобритании (Уэльс) и ранней юры (формация Кайента, синемюрский / плинсбахский ярус) США (штат Аризона).
Помимо географической удаленности, установленные виды отличаются особенностями зубочелюстного строения. В частности, у китайских форм в большей степени редуцированы коренные зубы. В то же время, некоторые наблюдаемые отличия могут объясняться возрастными изменениями. Особенно это справедливо в отношение количества зубов, особенно предкоренных, число которых выше у ювенильных особей. У взрослых особей Morganucodon watsoni I-й верхний предкоренной зачастую утрачивается, а последний коренной варьирует в размерах и строении. Кроме того, детали окклюзии коренных зубов у разных экземпляров Morganucodon watsoni несколько отличаются.

Пластическая реконструкция внешнего облика Morganucodon.
Буккальный цингулюм и промежутки между буграми верхних коренных намного лучше развиты у Morganucodon oehleri, чем у Morganucodon heikuopengensis и Morganucodon watsoni. Различие в высоте между бугром A и бугром B и развитие цингулярных бугорков является намного более явным у Morganucodon oehleri, чем у Morganucodon heikuopengensis. Кроме того, наибольший послеклыковой зуб — это M2 у Morganucodon oehleri и M1 у Morganucodon heikuopengensis, но последний предкоренной, P4, больше у Morganucodon watsoni. M3 намного меньше у Morganucodon heikuopengensis, чем у Morganucodon oehleri. Другие особенности включают величину верхнего бугра D и его топографическое соотнесение с бугром C. Также последний коренной у китайских Morganucodon heikuopengensis и Morganucodon oehleri намного меньше, чем e Morganucodon watsoni из Великобритании. В настоящее время невозможно установить, присущи ли китайским таксонам полиморфные различия, подобные имеющимся у Morganucodon watsoni. Учитывая все это, нельзя исключать возможность, что некоторые виды Morganucodon заслуживают выделения в самостоятельные рода.

Морганукодон (Morganucodon sp.) в естественном окружении. Автор — Х. Ласерда.
Образ жизни
Наличие мелких ямок на морде морганукодонов говорит о наличии вибрисс, а крупные глазницы — о сумеречной или ночной активности. Светлое время суток эти животные проводили в норах и других укрытиях. Строение зубов показывает, что они являлись скорее плотоядными, чем всеядными. Рацион мог состоять из насекомых и других беспозвоночных.
Размножение
Скорее всего, морганукодоны откладывали небольшие яйца в пергаментной оболочке. Благодаря находкам остатков их беззубых младенцев и молодых особей с единственным набором молочных зубов можно предполагать, что это были одни из первых млекопитающеобразных, детеныши которых полностью зависели от матери и ее молока на самых ранних этапах своей жизни. Судя по всему, молоко выделялось из участков кожи на брюхе.

Семейная группа Morganucodon преследуют клювоголовое Clevosaurus, похитившее одного из их детенышей. Автор — М. Уиттон.
Ссылки
Морганукодон (Википедия) перейти
Morganucodon (Paleobiology Database) перейти
Литература
Кэрролл Р. Палеонтология и эволюция позвоночных: В 3-х т. Т. 2. — М.: Мир, 1993
Россолимо О. Л., Павлинов И. Я., Крускоп С. В., Лисовский А. А., Спасская Н. Н., Борисенко А. В., Панютина А. А. Разнообразие млекопитающих. Ч. 1-3. — М.: Изд-во КМК, 2004
Татаринов Л. П. Эволюция териодонтов. — М.: Наука, 1976
Kielan-Jaworowska Z., Cifelli R.L., Luo Z.-X. Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure. — Columbia University Press, 2004