Носороги шерстистые — Coelodonta
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Клада Скротиферы (Scrotifera)
Клада Феревунгуляты (Fereuungulata)
Клада Эвунгуляты (Euungulata)
Клада Мезаксонии (Mesaxonia)
Отряд Непарнопалые (Perissodactyla)
Подотряд Тапирообразные (Tapiromorpha)
Инфраотряд Носорогообразные (Ceratomorpha)
Надемейство Риноцеротоиды (Rhinocerotoidea)
Семейство Носороговые (Rhinocerotidae)
Подсемейство Риноцеротины (Rhinocerotinae)
Триба Риноцеротини (Rhinocerotini)
Подтриба Дицерорина (Dicerorhinina)
Род †Носороги шерстистые (Coelodonta)
Вид †Тибетский шерстистый носорог (Coelodonta thibetana)
Вид †Нихэваньский шерстистый носорог (Coelodonta nihowanensis)
Вид †Тологойский шерстистый носорог (Coelodonta tologoijensis)
Вид †Обыкновенный шерстистый носорог (Coelodonta antiquitatis)
Носороги шерстистые (Coelodonta Bronn, 1831) — род подсемейства риноцеротин (Rhinocerotinae Gray, 1821), происходящий из позднего плиоцена - раннего голоцена Азии, среднего - позднего плейстоцена Европы и позднего плейстоцена Африки. Установлены 4 вида.

Реконструкция внешнего облика обыкновенного шерстистого носорога (Coelodonta antiquitatis). Автор — В. Симеооновски.
Синонимы: Rhinoceros Linnaeus, 1758 (частью); Gryphus Schubert, 1823, non Muhlfeldt, 1811; Hysterotherium Giebel, 1847; Tichorhinus Brandt, 1849; Atelodus Pomel, 1853 (частью); Coelorhinus Frech, 1904; Procerorhinus Kretzoi, 1942.
Родовое название представляет собой сочетание древнегреческих слов κοιλία (koilía) — "пещера, полость" и ὀδούς (odoús) — "зуб", означая в переводе "полый зуб", "полостнозуб". Оно указывает на наличие характерных лунок в щечных зубах животного.
Строение и функции
Шерстистые носороги имели крупные для семейства размеры: длина их тела достигала 3-4,3 м, высота в холке — 1,3-2 м.
Череп сильно удлинен. Носовые кости длинные, широкие, загнуты вниз. Есть костная носовая перегородка, у взрослых полная (у самок, возможно, не всегда). Подобно всем Dicerorhinina, имеется два рога, по одному на носовых и лобных костях. На местах их прикрепленя к черепу находятся хорошо выраженные шероховатости, более обширные, чем у других членов подтрибы. Носовой рог очень крупный, наклоненный вперед и сплюснутый с боков. Затылочный отдел черепа в меньшей или большей степени отклонен назад. Симфизный отдел нижней челюсти спереди расширен.
Зубная формула: I0/i0, C0/c0, P3/p3, M3/m3. Имеются молочные очень маленькие резцы, изредка закладываются постоянные у взрослых особей. Щечные зубы гипсодонтные. Их высокие коронки имеют призматическую форму и плоскую жевательную поверхность. Эмаль толстая, на боковых сторонах коронки грубо шероховатая, с цементом. Поперечные гребни на верхних коренных направлены очень косо к продольной оси зуба. Для зубов характерны колодцеобразные долинки с вертикальными стенками, в отличие от более древних носорогов, зубные долинки которых имеют вид пологих воронок. Криста и кроше хорошо развиты, не разветвлены, сливаются друг с другом при стирании зуба, отделяя дополнительную замкнутую долинку. В сравнении с зубами современных носорогов, щечные зубы более мощные, высокие и с утолщенной эмалью, больше всего напоминающие таковые белого носорога (Ceratotherium simum).
Туловище бочкообразное, с высоким загривком и укороченными, относительно массивными конечностями. Характерным признаком являлся хорошо развитый шерстный покров, защищавший от низких температур и холодных ветров. Низко посаженная голова с квадратными губами позволяла собирать основной корм — степную и тундростепную растительность.

Прорисовка реконструированного скелета обыкновенного шерстистого носорога (Coelodonta antiquitatis). Автор — М. Антон.
Состав
Тибетский шерстистый носорог (Coelodonta thibetana Deng, Wang, Fortelius, Li, Wang, Tseng, Takeuchi, Saylor, Säilä et Xie, 2011) — вид, известный из позднего плиоцена северного Китая.
Нихэваньский шерстистый носорог (Coelodonta nihowanensis Kahlke, 1967) — вид, известный из раннего плейстоцена северного Китая.
Тологойский шерстистый носорог (Coelodonta tologoijensis Beliajeva, 1966) — вид, известный из раннего - среднего плейстоцена северного Китая, Монголии, Забайкалья и Центральной Европы.
Обыкновенный шерстистый носорог (Coelodonta antiquitatis (Blumenbach, 1799)) — типовой вид, известный из среднего плейстоцена - раннего голоцена большей части Европы и Северной Азии, а также позднего плейстоцена крайнего севера Африки.
Эволюция
Ввиду эндемичного распространения рода Coelodonta на территории Евразии и общих черт строения изначально предполагалось более тесное родство его с азиатскими, чем с африканскими носорогами. Соответственно, рассматриваемый род был помещен в подтрибу Dicerorhinina. С анатомической точки зрения, род Coelodonta наиболее тесно связан с родом Stephanorhinus, известным начиная с конца позднего миоцена Западной Евразии. Они образуют общую кладу, именуемую группой Coelodonta-Stephanorhinus. Для обоих родов характерно окостенение носовой перегородки, среди Dicerorhinina встречающееся только у них. Первые признаки окостенения относительно рано фиксируются у Stephanorhinus и постепенно прогрессируют от лесных форм до степных, пока, наконец, не достигают максимумального выражения у обыкновенного шерстистого носорога (Coelodonta antiquitatis).
Еще одна общая морфологическая черта Coelodonta и Stephanorhinus — отсутствие резцов среди постоянных зубов, тогда как в молочной зубной системы они рудиментарны. С другой стороны, у их ближайшего современного родственника — почти вымершего суматранского носорога (Dicerorhinus sumatrensis) — присутствуют постоянные резцы, хотя и в сокращенном числе. Молекулярно-генетический анализ и изучение белковых последовательностей подтверждают тесную связь упомянутых трех родов. С помощью методов палеопротеомики удалось установить, что Stephanorhinus парафилетичен по отношению к Coelodonta. Судя по всему, последний произошел от ранней линии Stephanorhinus.
Родственные связи носорогов, включая шерстистых, представлены на следующей кладограмме:
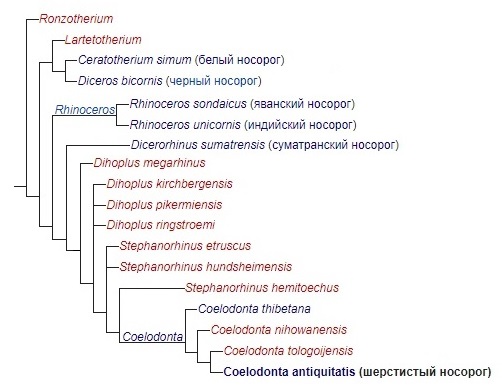
Вероятно, первоначальная эволюция рода Coelodonta происходила во влажных лесах, чем объясняется отсутствие его остатков миоценового возраста. Вервые данный род фиксируется в позднем плиоцене (около 3,7 млн. лет назад) Тибетского плоскогорья. Более 3 млн. лет эволюция шерстистых носорогов протекала на просторах Центральной Азии. Предполагается, что они могли населять весьма разнородные ландшафты — от хорошо увлажненных речных долин до иссушенных равнинных возвышенностей. Изначально рацион этих носорогов был довольно разнообразен и состоял главным образом из листьев нижнего яруса деревьев и кустарников, но включал также молодые побеги, небольшие веточки, мхи, различные травы и т. п.
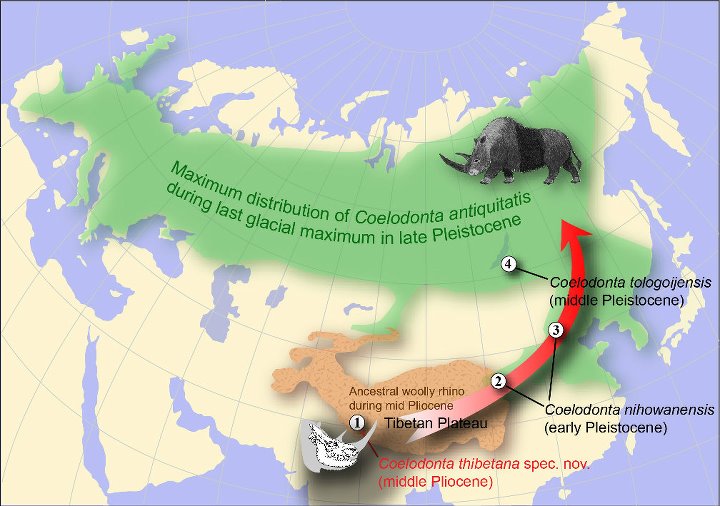
Распространение шерстистых носорогов (Coelodonta): 1 — тибетский шерстистый носорог (Coelodonta thibetana), 3,7 млн. лет назад; 2, 3 — ихэваньский шерстистый носорог (Coelodonta nihowanensis), 2,5 млн. лет назад; 4 — тологойский шерстистый носорог (Coelodonta tologoijensis), 0,75 млн. лет назад; зеленым цветом показана область максимального распространения самого известного и массового вида обыкновенный шерстистый носорог (Coelodonta antiquitatis) во время последнего ледникового максимума, около 27-20 тыс. лет назад.
На протяжении плейстоцена климат Земли претерпел значительные изменения. Постепенно увеличивались сезонные колебания температур: зима становилась прохладнее, а лето более засушливым. Это происходило на фоне всеобщего похолодания, способствующего накоплению в высоких широтах, а также в горных массивах громадных ледовых масс. Под влиянием прогрессирующего понижения среднегодовых температур и иссушения климата на обширных пространствах Северной Евразии леса и лесостепи отступали перед степями и приледниковыми тундростепями.
Некоторые тибетские виды, приспособившиеся к жизни в открытых ландшафтах, вероятно, смогли переселиться с высокогорных пастбищ на холодные равнины, раскинувшиеся к северу от их прародины. Среди них были и шерстистые носороги. Вслед за расширяющимися степями они сначала распространились к северу и западу от Китая, а в начале среднего плейстоцена проникли и в Европу, попутно замещая своих теплолюбивых предшественников из родов Stephanorhinus и Dihoplus. В период максимального распространения в конце позднего плейстоцена ареал шерстистых носорогов простирался от Западной Европы до Приморья.
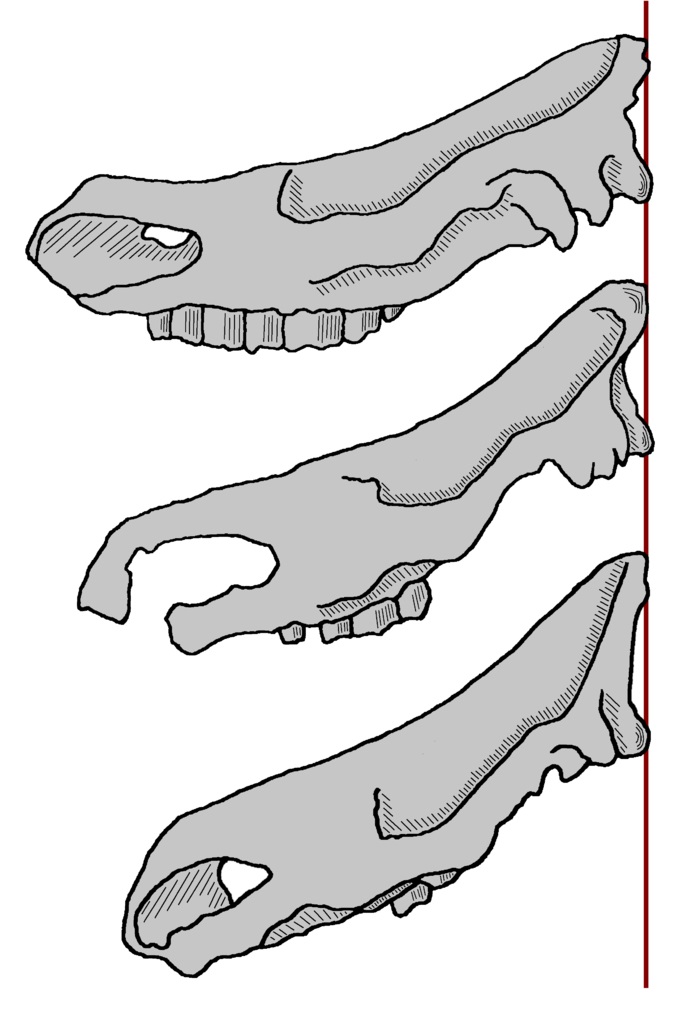
Схема, демонстрирующая степень наклона черепа различных представителей рода Coelodonta, сверху вниз: нихэваньский шерстистый носорог (Coelodonta nihowanensis), тологойский шерстистый носорог (Coelodonta tologoijensis) и обыкновенный шерстистый носорог (Coelodonta antiquitatis).
Усиливающийся континентальный климат наложил свой отпечаток на внешний облик и образ жизни этих животных, создав поистине северных носорогов — одних из самых эффективных потребителей жесткой травянистой растительности в холодных тундростепях и характернейших представителей мамонтовой фауны. Претерпела изменения их морфология.
Постепенно иным стало положение головы — она опустилась ниже к поверхности грунта. Лицевой отдел черепа удлинился и сузился, глазницы передвинулись ближе к затылку, а сама затылочная поверхность сильно отклонилась назад. Благодаря смещенным вверх глазам животное могло обозревать местность во время пастьбы с низко наклоненной головой. Пропорционально лицевому отделу удлиняется и мозговой отдел черепа, что связано с удлинением лобных и теменных костей. При этом высота черепа пропорционально уменьшается, а затылочный гребень увеличивается.
При увеличении линейных размеров черепа увеличивается массивность и толщина костей его составляющих, особенно теменных, лобных и носовых. Толщина внешнего слоя компакты теменной кости достигает у шерстистых носорогов 1,5-2,5 см. Обширные лобно-теменные синусы, разделенные многочисленными перегородками, создают пневматизацию массивных костей (подобно пневматизации костей мозгового черепа у слонов), что служит для некоторого облегчения черепа.
При пастьбе в зимние снежные периоды кожные ткани конца морды подвергались механическому раздражению. Это, в свою очередь, сильнее стимулировало рост филаментов носового рога, который в этих условиях также употреблялся для разгребания снега. Для более эффективного выполнения этой задачи носовой рог увеличился, наклонился вперед и стал плоским с боков. Носовые кости удлинялись, а их передние концы все больше загибались книзу, стремясь к предчелюстным костям и в конечном итоге сраслись с ними. Общая площадь носовых костей значительно увеличилась, создав условия для более прочного крепления носового рога, который достигал больших размеров. Увеличившиеся силы давления основания массивного рога на носовые кости стимулировали полное окостенение носовой перегородки.

Продольный разрез черепа обыкновенного шерстистого носорога (Coelodonta antiquitatis).
Из наблюдений в природе за современными носорогами известно, что они активно употребляют свой носовой рог для обороны от хищников, для защиты кормовой территории от особей своего вида, для рыхления земли у высохшего дна русла реки при добыче воды, для корчевки кустарника и раздвигания его плотных зарослей. Однако всех этих функций, выполняемых носовым рогом, оказалось недостаточным, чтобы закрепить у современных видов носорогов мутации прижизненного окостенения их носовой перегородки. На протяжении всей их жизни она остается хрящевой. И у носорогов из неогеновых отложений носовая перегородка не сохранилась, что может указывать на ее хрящевую природу при жизни животных, и косвенно свидетельствовать о жизни их в тропическом климате, как и современных видов носорогов.
Только у плейстоценовых носорогов родов Coelodonta, Stephanororhinus, Dihoplus и Elasmotherium мутация, приведшая к прижизненному окостенению носовой перегородки, закрепилась в условиях глобального похолодания климата. Однако у родов Stephanororhinus и Dihoplus она не достигла максимального развития окостенения, как это наблюдается у родов Coelodonta и Elasmotherium. Этот факт еще раз подтверждает, что и носороги рода Elasmotherium, которые сформировались в одно время (плиоцен) с носорогами рода Coelodonta, и на одной территории (север Китая, Монголия), находились в момент своей эволюции в сходных условиях — резко континентальном климате, с низкими зимними температурами, и хотя бы минимальным снежным покровом.
О активном употреблении шерстистыми носорогами носового рога для добывания корма из-под снега в длительный зимний период указывают характерные потертости на сохранившихся в мерзлой почве рогах и особенно сильное развитие костных экзостозов (мелких шипообразных костных выростов) на носовых и лобных костях. Они служили дополнительной защитой кровеносным сосудам, питающим рог, от сильного сжатия, когда животное активно работало рогом. Лобный рог не участвовал в работе, но при большом переднем роге, тоже достигал значительных размеров, так как стимулировался к росту мощно развитой кровеносной сетью питавшей "рабочий" рог.
Значительных размеров диастемное (беззубое) пространство резцовых костей, располагающееся перед рядом щечных зубов, указывает на большую вместительную ротовую полость и хорошо развитую мускулатуру верхней и нижней губы. Это особенно имело большое значение для согревания холодной пищевой массы, которая поступала в ротовую полость частично смешанная со снегом в зимнее время. Щечные зубы эволюционировали в сторону питания сухой травой: возросла высота коронок, увеличилась доля отлагаемого на них цемента. Вместе с лицевым отделом черепа носовые дыхательные ходы удлинились и стали уже, способствуя согреванию вдыхаемого зимой воздуха. Удлинение костного неба и отодвигание хоан привело к увеличению объема ротовой полости, что важно было для согревания пищевой массы смешанной со снегом. Увеличились общие размеры животных, укоротились и стали более массивными их конечности. Для защиты от усиливающихся холодов развился плотный волосяной покров.

Обыкновенный шерстистый носорог (Coelodonta antiquitatis), добывающий траву из-под снега.
В конце плейстоцена - начале голоцена шерстистые носороги повсеместно исчезают. Предполагается, что это произошло в основном из-за климатических изменений, сопровождавших окончание последней ледниковой эпохи — вследствие потепления и увеличения влажности могла значительно уменьшиться площадь подходящих местообитаний.
Ссылки
Марков А. Шерстистые носороги пришли с Тибета. — 2011 (Элементы.Ру) перейти
Coelodonta (Paleobiology Database) перейти
Coelodonta (Википедия) перейти
Литература
Гарутт Н. В. Шерстистый носорог (Морфология, систематика, геологическое значение). — С.-П., 1998
Основы палеонтологии. В 15 т. Т. 13: Млекопитающие / Гл. ред. Орлов Ю. А. — М.: Государственное научно-техническое издательство литературы по геологии и охране недр, 1962
Deng T., Wang X., Fortelius M., Li Q., Wang Y., Tseng Z. J., Takeuchi G. T., Saylor J. E., Säilä L. K., Xie G. Out of Tibet: Pliocene woolly rhino suggests high-plateau origin of Ice Age megaherbivores. Science 333, 2011
Kahlke H.-D. Objevy ve čtyřech světadílech. — Praha: Orbis, 1973