Строение и функции
В настоящее время пума — четвертый по величине представитель семейства кошачьих в мире и второй в Америке. Крупнее нее только тигр, лев и ягуар, а пятое место занимает леопард. Пума является второй по величине американской кошкой после ягуара. Длина ее головы и туловища составляет 105-196 см, длина хвоста — 67-78 см, высота в холке — 63-76 см. Масса варьирует в пределах 36-125 кг; самки мельче самцов приблизительно на 20-30 %. Разные подвиды сильно отличаются по размеру: самые мелкие обитают в тропиках, наиболее крупные — в северных и южных частях ареала. Обычно нормальный самец крупного подвида весит 60-80 кг. Самая крупная зарегистрированная особь происходит из Аризоны.

Пума (Puma concolor) в зимнее время.
Голова относительно небольшая, несколько вытянутая. Цвет радужки глаз варьирует от серовато-коричневого до золотистого; зрачки овальные. Уши маленькие, округлые. Телосложение очень мощное. Но при этом подтянутое. Туловище удлиненное, сильное и гибкое. Конечности мускулистые. Их задняя пара заметно длиннее и массивнее передней, благодаря чему пума отлично прыгает. Лапы широкие и мощные. На кисти 4, на стопе 5 пальцев с кривыми и острыми втяжными когтями. Когти используются для захвата и удержания добычи, а также для лазания по деревьям. Подушечки пальцев овальные, на подушечке пятки три отчетливых доли — общая черта для всех кошек. Хвост длинный и упругий, равномерно опушенный.

Портрет пумы (Puma concolor).
Мех на теле сравнительно короткий, грубый и густой. Его окрас однотонный, серовато-коричневый или коричневато-желтый; нижняя сторона тела светлее верхней. На груди, горле и брюхе имеются белесые подпалины, на морде — черные отметины, конец морды белый, уши темные, кончик хвоста черный. Цвет шерсти зависит от места обитания: у животных из тропических районов он более насыщенный, с рыжеватым оттенком, тогда как у северных — более бледный, сероватых тонов. Для особей из Анд характерен серебристо-голубоватый окрас. Также известно о существовании светлых и даже белых пум, а также темно-коричневых, встречающихся преимущественно в Латинской Америке. Пумы-альбиносы и меланисты в природе неизвестны. Наряду с ягуарунди пума — единственная одноцветно окрашенная американская кошка. В целом, окрас пумы напоминает таковой ее основной добычи — оленей. У горных и северных подвидов зимний мех заметно длиннее летнего.
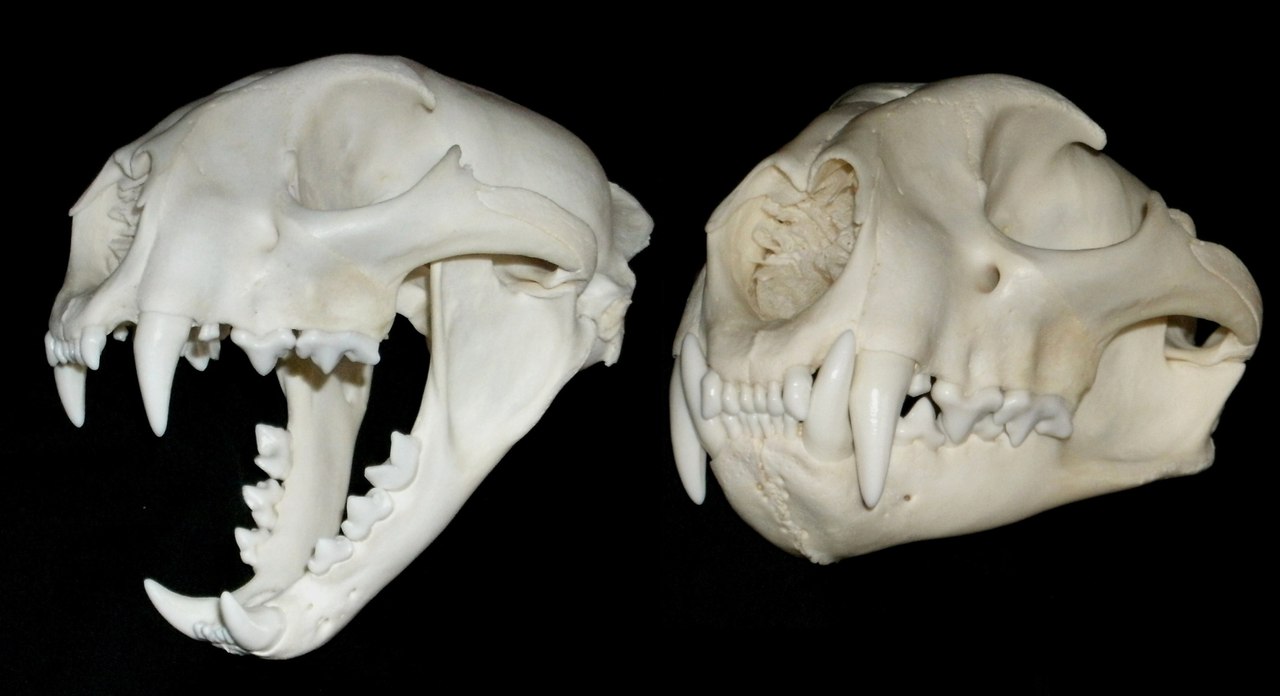
Череп с нижней челюстью пумы (Puma cocncolor).
Череп по пропорциям похож не на черепа кошек рода Panthera, а на увеличенный вариант черепа мелкого кошачьего, но с более массивными челюстями и скуловыми дугами, а также сильнее выраженными гребнями и отростками. Крыша черепа куполообразная. Граница между полостями слухового барабана практически не прослеживается. Наружная барабанная полость относительно мала, что является характерной особенностью Puma concolor.
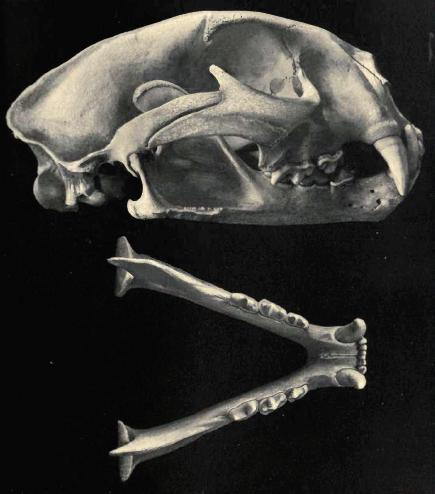
Череп и нижняя челюсть пумы (Puma cocncolor).
Сбоку нижней челюсти находятся два крупных подбородочных отверстия (одно расположено ниже дальней границы клыка, а другое ниже заднего корня р3) и одно маленькое выше среднего отверстия. Ямка большой жевательной мышцы достигает уровня протоконида m1. Суставная поверхность нижнечелюстного мыщелка верхне-наружно сильно закруглена. Среднее значение индекса массивности нижней челюсти (высота тела челюсти за m1 × средне-дальняя длина m1 × 100) — 146,2.
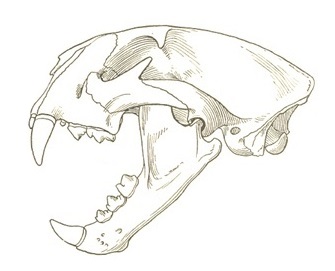
Прорисовка черепа с нижней челюстью пумы (Puma cocncolor).
Зубная формула: I3/i3, C1/c1, P3/p2, М1/m1 = 30. I3 имеет два небольших бугорка, один дальний и один среднеязычный. Р2 очень маленький. P3 является относительно просто устроенным зубом с двумя бугорками — более крупным основным и меньшим следующим за ним, а также задне расположенным цингулюмом. Снаружи P3 имеет смещенный вперед край, который несет небольшой бугорок, подверженный индивидуальной изменчивости. P4 с редуцированным протоконом; степень развития эктопарастиля варьирует от особи к особи.
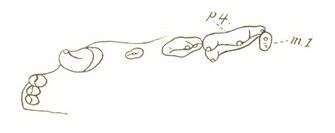
Прорисовка левой стороны верхней челюсти пумы (Puma cocncolor).
За нижним клыком расположена довольно широкая диастема. р3 имеет небольшой цингулюм и хорошо развитый, закругленный передний дополнительный бугорок, выступающий сильнее, чем дополнительный задний. р4 несет симметричный протоконид (при взгляде снаружи) и хорошо развитыи дополнительные бугорки. Протоконид этого зуба узкий и симметричный (при взгляде снаружи). Дальний дополнительный бугорок, выступающий и немного сдавленный с внешней стороны, несет хорошо развитый дальний цингулюм. m1 мощный, с вертикально ориентированным протоконидом, который чуть длиннее и выше, чем параконид; оба бугорка ориентированы вертикально.
По сравнению с представителями родом пантер (Panthera), клыки и хищнические зубы у пумы (Puma concolor) сравнительно небольшие. У ягуара (Panthera onca) клыки более крупные, но p3 меньше и m1 длиннее. В сравнении с небольшими кошками — такими, как ягуарунди (Herpailurus yagouaroundi) — клыки пумы более круглые в сечении, а P3 длиннее. По сравнению с близко родственным вымершим мирацинониксом (Miracinonyx trumani), у пумы относительно более крупные клыки, меньший P3, более редуцированный протокон на P4, меньший p4 и больший m1.
Состояние зубов — один из основных показателей при определении возраста пумы. Молочные зубы у нее полностью прорезаются к 4 месяцам жизни. Постоянные зубы начинают прорезаться в 6-8 месяцев, и к возрасту в 1,5-2 года прорезаются полностью. С возрастом клыки и резцы сильно стачиваются и темнеют.
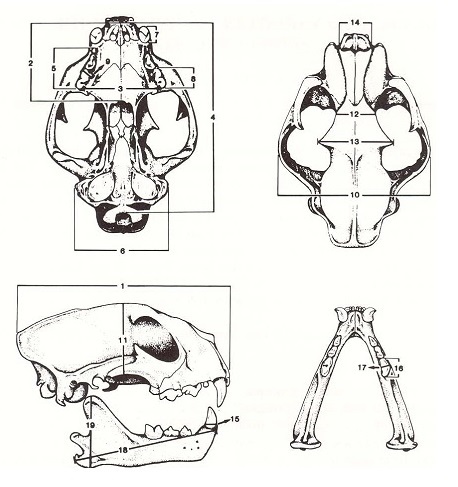
Черепные, нижнечелюстные и зубные показатели, варьирующие у различных подвидов пумы (Puma cocncolor): 1) наибольшая длина черепа; 2) небная длина; 3) межчелюстная ширина; 4) основная длина; 5) длина альвеолярной линии; 6) ширина сосцевидного отростка; 7) наибольший диаметр верхнего клыка; 8) наибольшая длина Р3; 9) наибольшая ширина Р3; 10) скуловая ширина; 11) глубина черепа; 12) межглазничная ширина; 13) надглазничная ширина; 14) носовая ширина; 15) наибольший диаметр нижнего клыка; 16) наибольшая длина m1; 17) наибольшая ширина m1; 18) наибольшая длина нижней челюсти; 19) глубина нижней челюсти.
Рост черепа продолжается на протяжении большей части жизни животного. Самцы продолжают расти до 7-9 лет, самки — до 5-6 лет. Череп ювенильных особей (приблизительно до 1 года) более гладкий и с менее развитым лицевым отделом и нижней челюстью. Имеется лишь слабый намек на заглазничные отростки и сагиттальный гребень, которые с возрастом становятся более рельефными. По мере взросления кости утолщаются, а верхнечелюстная и нижнечелюстная области увеличиваются. Молочные зубы сменяются постоянными примерно к 2 годам.
Коэффициент длины поясничной области позвоночника к грудной составляет приблизительно 0,78. Невральные дуги поясничных позвонков расширены передне-задне. Длина хвоста составляет 90 % длины прекаудального позвоночного столба.


Смонтированный скелет пумы (Puma cocncolor).
Акромион в верхней части лопатки выступает над суставной ямкой. Ближняя часть стержня плечевой кости сжата срединно-наружно, дельтопекторальный гребень небольшой, наружный гребень для крепления плечелучевой мышцы уменьшен. Также плечевая кость имеет небольшой, но хорошо развитый срединный надмыщелок и глубокую локтевую ямку. Ямка лучевой кости широкая и неглубокая, короновидная ямка овальная и глубокая. Лучевая кость составляет почти 90 % длины плечевой кости. Ближняя суставная поверхность ориентирована под небольшим углом к стержню в дальней проекции, суставная фасетка для соединения с локтевой костью имеет эллиптическую форму. Ее длинная ось ориентирована перпендикулярно к стержню. Запястные суставы относительно крупные, дальняя суставная поверхность трапецоида разделена гребнем. Ближние и дальние концы плюсневых костей утолщены.
Длина бедренной кости составляет 22,6-27,4 см. Большая берцовая кость имеет тонкое тело, отношение между минимальной длиной его окружности и длиной кости — 0,234 +/- 0,006. Ближняя часть плантарной срединной фасетки сжатасрединно расширенной пазухой предплюсны. Палмарная наружная фасетка срединно-наружно сжата, с верхней стороны шейка относительно длинная. Стержень бедра выглядит прямым в сагиттальном плане. Коленная чашечка имеет слабовыпуклую поверхность соединения с бедром. Средний гребень большеберцового блока вытянут назад при взгляде снизу; у ягуара этот выступ является широким и тупым.
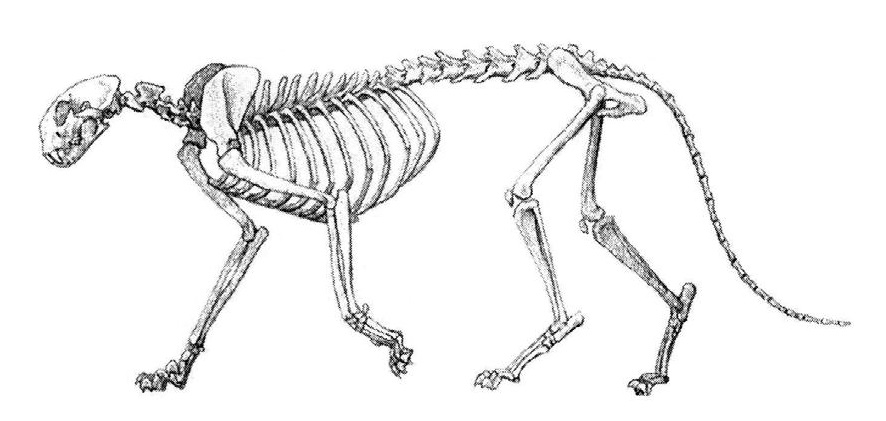
Прорисовка скелета пумы (Puma cocncolor) в естественной позе.
Отношение голени к длине бедра — приблизительно 0,90. В дальней проекции суставная поверхность иззубрена вдоль заднего края. У таранной кости имеется ладьевидная суставная поверхность, пролегающая срединно. Шейка таранной кости относительно более длинная, чем у ягуара (Panthera onca). Средняя сторона кости несет мелкое и широкое вертикальное углубление, в отличие почти плоской поверхности у ягуара. Средняя пяточная поверхность округлая, с небольшим ближним расширением. Боковая пяточная суставная поверхность выглядит менее вогнутой, чем у гепарда (Acinonyx jubatus). В суставном сочленении таранная кость почти соприкасается с задним краем пяточной кости.
Ближняя суставная поверхность трапецевидной кости является вогнутой, суставная поверхность клиновидной кости имеет яйцевидную форму. Пястные кости с хорошо развитыми выступами мускульных прикреплений — например, для разгибающих мышц запястий. Также пума имеет общую с Miracinonyx и современной ягуарунди (Herpailurus yagourundi) форму ближнего конца IV-й пястной кости с заметным в передней проекции зубцом. В ладьевидной кости умеренно развита бугристость, расширяющая ее ближне больше, чем у современного Acinonyx. В этом отношении она аналогична таковой рода Miracinonyx. Фасетка ладьевидной кости для соединения с крючковидной не настолько сдвинута вниз, как у Miracinonyx, и является почти вертикальной. Также наличествует небольшая, вертикально ориентированная поверхность для сочленения с пяточной костью.
Ближняя поверхность кубовидной кости выглядит слабовыпуклой и имеет маленькую поверхность для сочленения с ладьевидной костью. В периферии кубовидная кость аналогична таковой у Miracinonyx и Acinonyx в наличии двух фасеток: большей для IV-й плюсневой кости и гораздо меньшей для V-й плюсневой кости. Однако плюсневые кости демонстрируют тенденцию к большему дальнему расширению по отношению к средней ширине костного стержня.
Генетика
В кариотипе пумы 38 хромосом.
Хотя пума не является близким родственником других ныне живущих кошек, в неволе были получены ее гибриды с леопардом, ягуаром и оцелотом. Гибрид пумы и леопарда (пумапард) склонен к карликовости и вдвое мельче родительских видов. Строением тела он похож на пуму. Его ноги короткие, а хвост длинный. Известны особи разной окраски: песчаной, темно-желтой или серо-коричневой. На шкуре есть пятна, как у леопарда, но не такие яркие.

Чучело пумпарда.
Эволюция и родственные связи
Ископаемые остатки вида Puma concolor известны начиная с раннего плейстоцена по голоцен. Они обнаружены в США (штаты Вашингтон, Айдахо, Монтана, Вайоминг, Юта, Калифорния, Аризона, Нью-Мексико, Техас, Канзас, Миссури, Огайо, Виргиния, Джорджия, Южная Каролина, Флорида), Мексике (штаты Халиско, Чиуауа), Панаме, Венесуэле (штат Гуарико), Перу, Боливии (департамент Тариха), Бразилии (штат Баия), Уругвае, Аргентине (провинции Буэнос-Айрес, Сантьяго-дель-Эстеро) Чили (область Магальянес).
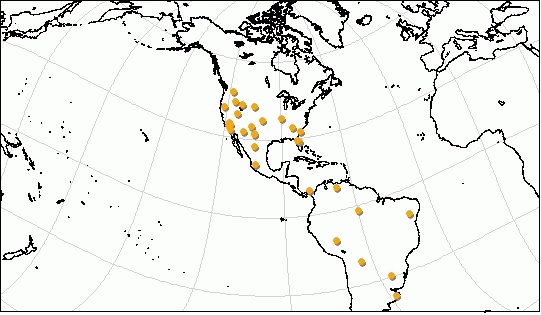
Некоторые местонахождения ископаемых остатков пумы (Puma cocncolor).
Наиболее полные остатки происходят из местонахождений позднеплейстоценового возраста Ранчо ла-Бреа и Мак-Китрик в Калифорнии, а также из Флориды, где Puma concolor столь же многочисленна, как и Panthera onca. Окаменелости из других мест обычно фрагментарны. Однако найденный материал доказывает, что географическое распределение плейстоценовой пумы практически совпадает с ее современным ареалом. Особи из ряда древних популяций были гораздо крупнее современных. Так, рассчеты предполагают, что пумы из плейстоцена Патагонии могли весить примерно до 145 кг.
Долгое время считалось, что род Puma сформировалcя в плейстоценовое время на территории Северной Америки. К примеру, отсюда известна относительно мелкая Puma lacustris, которая, возможно, представляет собой раннюю форму Puma concolor (Репеннинг с соавт., 1995 г.). Однако на евразийском материке в позднем плиоцене - среднем плейстоцене тоже обитала пумообразная кошка — Puma pardoides, также известная как Panthera (Viretailurus) schaubi. Их статки морфологически почти сопоставимы с соответствующими элементами скелета современной Puma concolor за исключением лишь более крупных размеров (Мадурель-Малапейра с соавт., 2010 г.).

Дистальный отдел конечности пумы (Puma cocncolor) из позднего плейстоцена Аргентины.
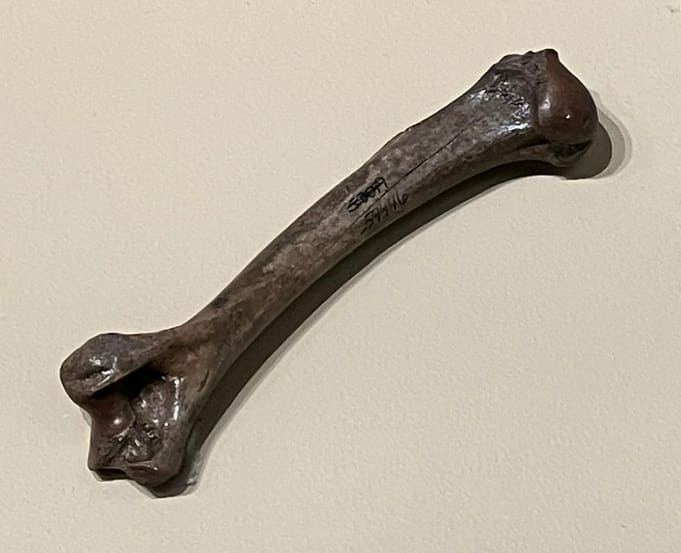
Плечевая кость пумы (Puma cocncolor) голоценового возраста, найденная в песчаных отмелях реки Канзас, США.
Находки Puma pardoides встречаются от Западной Европы до Центральной Азии во временном интервале от позднего плиоцена до конца раннего плейстоцена, совпадая по времени с появлением леопарда. Это подразумевает, что если Puma pardoides и современная Puma concolor являются родственными таксонами, предки последней должны была проникнуть в Америку по Берингийскому перешейку в самом конце плиоцена. В связи с этим необходима более тщательная ревизия азиатских окаменелостей для того, чтобы разобраться — являются ли они остатками пумообразных кошек, неверно идентифицированными по причине их сходства с остатками леопарда и снежного барса. Помимо этого, на роль предковой формы для линии пумы помимо европейской Panthera (Viretailurus) schaubi (Куртен, 1976 г.; Сотникова, 1978 г.) предполагались еще две кошки Старого Света — африканская Panthera crassidens и азиатский Metailurus (Петер и Хауэлл, 1976 г.).
Зубы пумообразных кошек, находимые в плейстоценовых отложениях на востоке Северной Америки, обычно упоминались в качестве Felis inexpectata (Коп, 1899). Данный автор даже определял эту форму в род Uncia и описывал ее как Uncia inexpectata, основываясь на материале, состоящем из нижней челюсти, ряда изолированных зубов и элементов скелета. По предположению Копа, Uncia inexpectata выглядел как крупная разновидность Uncia, "столь же большая, как ягуар".
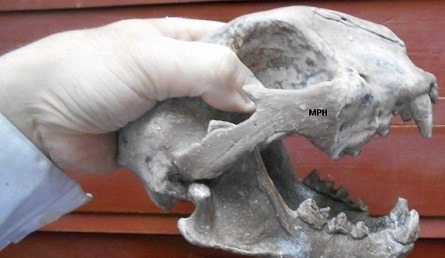
Череп с нижней челюстью пумы (Puma cocncolor) из голоцена Аргентины.
Более поздние исследования показали, что Felis inexpectata вполне сопоставима по размерам с современными крупными подвидами пумы Рuma concolor hippolestes и Рuma concolor oregonensis (Симпсон, 1941 г.). Чуть раньше Симпсона это предположение высказали Гидли и Гэзин (1938 г.), основываясь на остатках эволюционно продвинутой кошки из Камберлендской пещеры в Мэрилэнде. Относительно неглубокая челюсть, переднее расположение ямки большой жевательной мышцы и ее заднее расположение за m1 и, особенно, небольшая величина p4 относительно m1 явно напоминает пуму и фактически исключает связь с ягуарами. Указанные образцы обнаружены поблизости от типового местонахождения Felis inexpectata и относятся к одной и той же геологической формации. Согласно этим данным, весьма вероятно, что форма Felis inexpectata все же конспецифична пуме и что большинство образцов на самом деле принадлежит пумам.
Аналогичная синонимия проведена и с Felis longicrus (Браун, 1908 г.) — таксону, сходному с Felis inexpectata, который был обнаружен в Конкордском ущелье Арканзаса и сперва отнесен к этой форме. Чуть позднее Браун выделил найденные остатки в отдельный вид, основываясь на массивности P4 и более крупном его протоконе, расположенным дальше и под более тупым углом. Согласно Гидли и Гэзину, Felis longicrus является подвидом или даже примером индивидуальной измененчивости Рuma concolor.

Следовые отпечатки пумы (Puma cocncolor) и гуанако (Lama guanicoe) из позднего плейстоцена Аргентины.
Кроме этих восточных образцов, явно относящихся к одной группе, различные остатки пумаобразных кошек, причем в ряде случаев относительно полные и превосходно сохранившиеся, были найдены возле Тихоокеанского побережья. На их основании были установлены три вида — Felis daagetti, Felis hawveri и Felis bituminosa. Детальное изучение показало, что имеющиеся данные не подтверждают таксономического различия между этими формами и что явное сходство с современным западным материалом позволяет считать данные образцы примерами внутривидовой изменчивости или полового диморфизма Рuma concolor (Симпсон, 1941 г.). В дальнейшем Рэй (1958 г.) и Куртен (1965 г.) подтвердили, что имеющийся материал по промерам и особенностям строения не отличается от соответствующих элементов скелета ныне живущей пумы.
В то же время Ван Валькенбург с соавт. (1990 г.) отнесли больший ряд фрагментов Felis inexpectata наряду с родственным видом Аcinonyx trumani к роду Miracinonyx и предположили, что он может входить в число близких родственников пумы. В этот род также включили Felis longicrus и Smilodontopsis mooreheadi, ставшие субъективными синонимами Miracinonyx inexpectatus. На то, что Miracinonyx и Puma являются сестринскими таксонами, указывают и более ранние работы (Сэведж, 1960 г.; Куртен, 1976 г.). Так, Харрингтон (1986 г.) приводит доводы, основанные на общих особенностях строения малой берцовой кости: гребень на ней присутствовует только у пумы, ягуарунди и мирациноникса. Аналогичная ситуация у этих кошек наблюдается и в отношении молоточка среднего уха.

Поздний плейстоцен Боливии. Пума (Puma cocncolor) охотится на макраухений (Macrauchenia patachonica). Можно предположить, что их молодые особи были привычной и легкой добычей для этого хищника. Автор — М. Антон.
В ходе исследований ДНК ягуарунди, пумы и ягуара наибольшее сходство было выявлено между ягуарунди и пумой (17 и 13 аллелей соответственно) по сравнению с генетическим материалом домашней кошки, в то время как ягуар имеет только 12 аллелей. Кроме того, данные исследования показали, что несмотря на то, что эти виды находятся под угрозой исчезновения, имеющийся материал все еще демонстрирует достаточный уровень генетического разнообразия (Морено с соавт., 2006 г.).
Ископаемые остатки Puma concolor подтверждают, что начиная с раннего плейстоцена она была широко распространена в границах современного ареала (Морган с соавт., 1997 г.). Если раньше доказывали, что Miracinonyx и Puma concolor могли иметь общего предка, жившего в Америке в позднем плиоцене (Джонсон с соавт., 2006 г.), предполагая американское происхождение рода Puma, то сегодня присутствие примитивных пум в позднем плиоцене Евразии свидетельствует в пользу евразийско-африканского происхождения группы гепардоподобных кошек Acinonyx - Miracinonyx - Puma, с раздельным распространением последних двух в Америке (Христиансен с соавт., 2009). Скорее всего, род Puma возник в Евразии, откуда и заселил американский материк, исчезнув затем на своей исторической родине.

Поздний плейстоцен США. Пума (Puma concolor) с добытым вапити (Cervus canadensis) и американский мастодонт (Mammut americanum). Автор — В. Симеоновски.
По имеющимся данным, в конце плейстоцена в Северной Америке вид Puma concolor вымер вместе с многими представителями мегафауны. Однако он выжил в Южной Америке и повторно заселил Северную Америку примерно 10 тыс. лет назад.