Офиакодоны — Ophiacodon
Класс †Синапсиды (Synapsida)
Подкласс †Пеликозавры (Pelycosauria)
Надотряд †Офиакообразные (Ophiacomorpha)
Отряд †Офиакодонты (Ophiacodontia)
Семейство †Офиакодонтовые (Ophiacodontidae)
Род †Офиакодоны (Ophiacodon)
Вид †Ophiacodon mirus
Вид †Ophiacodon navajovicus
Вид †Ophiacodon uniformis
Вид †Ophiacodon retroversus
Вид †Ophiacodon hilli
Вид †Ophiacodon major
Офиакодоны (Ophiacodon Marsh, 1878) — род семейства офиакодонтовых (Ophiacodontidae Nopsca, 1923), происходящий из позднего карбона - ранней перми Северной Америки и, возможно, ранней перми Европы. Установлены 6-8 видов.

Реконструкция офиакодона (Ophiacodon mirus) с добытым антракозавром арчерией (Archeria crassidiscus). Автор — Д. Богданов.
Синонимы: Clepsydrops Cope, 1875 (частью); Theropleura Cope, 1878 (частью); Dimetrodon Cope, 1878 (частью); Diopeus Cope, 1892; Poliosaurus Case, 1907; Arribasaurus Williston, 1914; Diapaeus Williston, 1916; Therosaurus Huene, 1925; Winfieldia Romer, 1925.
История изучения
Данный род был описан американским палеонтологом Отниелем Чарлзом Маршем в 1878 г. Его название переводится как "змеиный зуб". Офиакодоны являются наиболее известными представителями своего семейства.
Строение и функции
Размеры их средние и крупные, вплоть до наибольших в семействе. Общая длина тела варьирует в пределах примерно от 1,2 до 4 м и более.
Череп длинный, узкий и высокий, очень большой относительно тела. Интересная особенность — наличие у некоторых крупных особей двух височных окон, описан также экземпляр с двумя окнами лишь на одной стороне (с другой было одно окно). Строение черепа предполагает наличие мощных челюстных мышц, способных осуществлять сильные укусы. Края челюстей несут множество зубов, представляющих собой острые сжатые конусы с режущими краями. Клыковидные зубы мало выделяются среди прочих зубов, стенки их альвеол не расширены. У более поздних офиакодонов дифференциация зубного ряда выражена несколько отчетливее, чем у ранних.
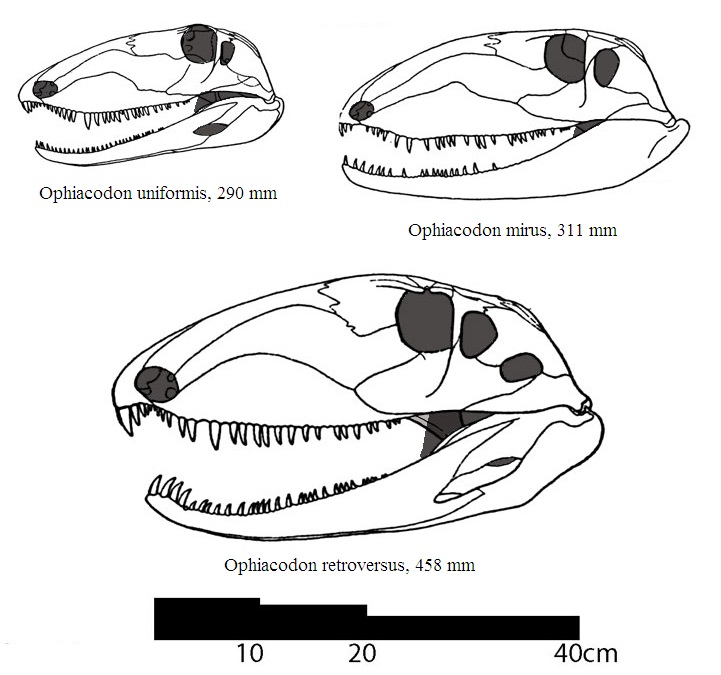
Прорисовки реконструированных черепов с нижними челюстями различных видов Ophiacodon с соблюдением общего масштаба.
Туловище у крупных видов массивное. Позвонки построены более легко, чем у мелких представителей семейства. Остевые отростки позвонков относительно высокие, но над кожей не выступавшие. Конечности короткие, со слабо окостеневшими суставами. Аддукторный гребень бедренной кости иногда редуцирован. Дистальные фаланги уплощенные, когти могли быть тупыми или даже отсутствовать.
Скелет офиакодонов демонстрирует интенсивный рост, вызываемый фиброламеллярной костной тканью, что может предполагать, по меньшей мере, их частичную гомойотермию. Такой тип костной ткани также распространен у птиц и млекопитающих.
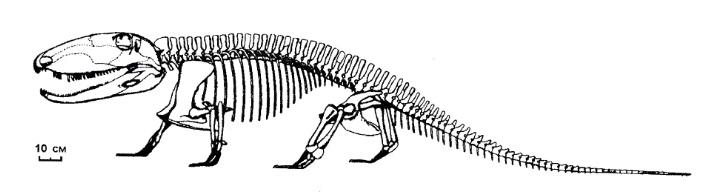
Прорисовка реконструированного скелета офиакодона (Ophiacodon retroversus).
Найденные остатки офиакодонов сильно различаются по своим размерам. Эти различия в прошлом часто использовались для разделения видов, однако сейчас они признаются лишь онтогенетической вариабельностью, т. е. связываются с различным возрастом отдельных животных. Меньшие кости часто имеют более слабо развитые суставные поверхности, чем более крупные кости, что подразумевает их принадлежность молодым особям, тогда как более крупные кости принадлежат взрослым. Гистологический анализ костей также свидетельствует о том, что различия в размерах зачастую объясняется различными стадиями индивидуального развития, а не свидетельствует о принадлежности к разным видам.
Признаваемые ныне виды офиакодонов очень близки между собой морфологически и отличаются в основном по размерам, распространению и геологическому возрасту. Лучше всего изученными являются Ophiacodon mirus, Ophiacodon uniformis и Ophiacodon retroversus.
В раннепермских отложениях Техаса рядом с неполным скелетом офиакодона было найдено яйцо длиной около 7,5 см, не успевшее подвергнуться развитию и сохранившееся в почти неповреждленном виде. Возможно, оно принадлежало этим животным.
Состав
Ophiacodon mirus Marsh, 1878 (тип) — типовой вид, известный из позднего карбона (гжельский ярус) - ранней перми (сакмарский / ? артинский ярус, приуральский отдел) штатов Нью-Мексико и Оклахома, США. Наличествует неполный скелет. Размеры некрупные, общая длина тела — около 1,6 м.



Реконструированный скелет офиакодона (Ophiacodon mirus) из ранней перми штата Нью-Мексико, США. Музей естествознания им. Филда, Чикаго.
Ophiacodon navajovicus (Case, 1907) Romer et Price, 1940 — вид, известный из позднего карбона (гжельский ярус) - ранней перми (ассельский / ? сакмарский ярус, приуральский отдел) штатов Нью-Мексико и Юта, США. Синонимы: Dimetrodon navajovicus Case, 1907; Arribasaurus navajovicus (Case, 1907) Williston, 1914. Известен лишь посткраниальный материал.
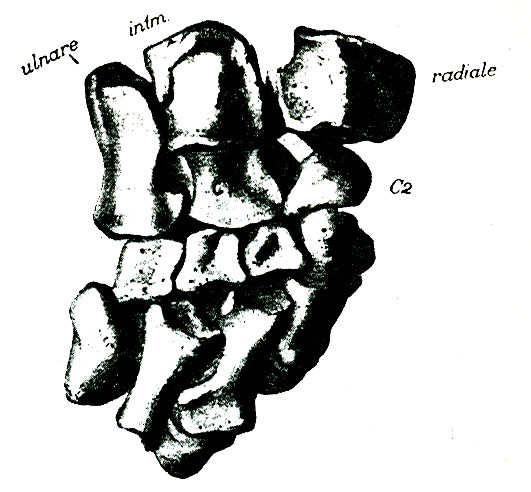
Прорисовка костей дистальной части передней конечности офиакодона (Ophiacodon navajovicus) из позднего карбона штата Нью-Мексико, США.
Ophiacodon uniformis (Cope, 1878) Romer et Price, 1940 — вид, известный из ранней перми (? ассельский / сакмарский - артинский ярусы, приуральский отдел) штатов Огайо, Оклахома и Техас, США. Синонимы: Theropleura uniformis Cope, 1878; Poliosaurus uniformis Case, 1907. Известен неполный скелет. Размеры мелкие, общая длина тела составляет порядка 1,5 м.


Реконструированный скелет офиакодона (Ophiacodon uniformis) из ранней перми штата Техас, США.
Ophiacodon retroversus (Cope, 1878) Romer et Price, 1940 — вид, известный из ранней перми (ассельский - артинский ярусы, приуральский отдел) штатов Оклахома и Техас, США. Синонимы: Theropleura retroversus Cope, 1878; Theropleura trianagulata Cope, 1878; Ophiacodon triangulata Cope, 1878; Clepsydrops leptocephalus Cope, 1884; Diopeus leptocephalus Cope, 1884; Therosaurus watsoni Huene, 1925. Вид известен по неполному скелету. Размеры крупные, общая длина тела — около 2,7 м.


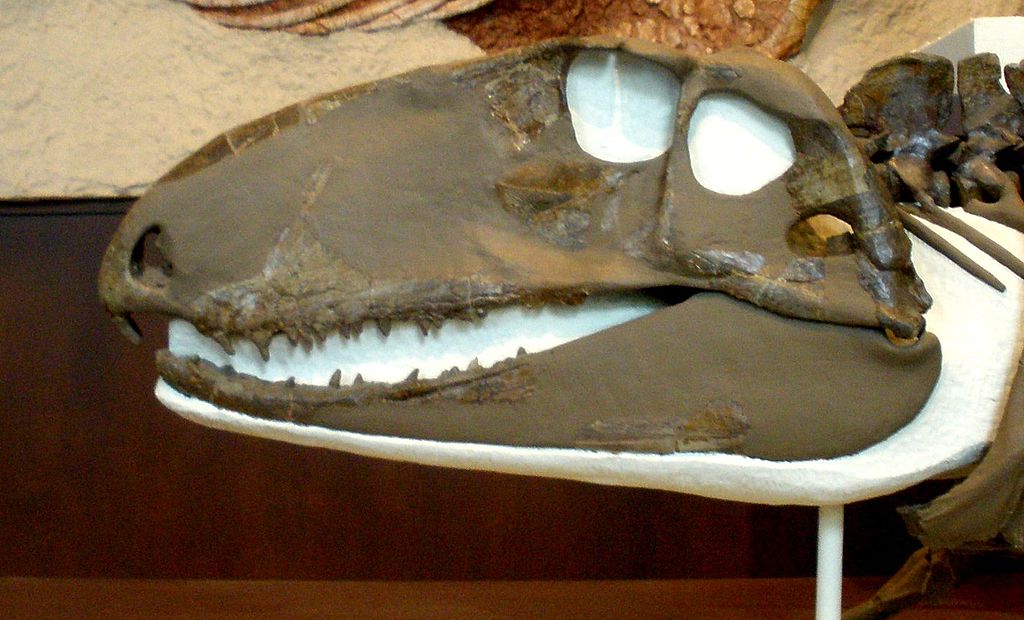

Реконструированный скелет офиакодона (Ophiacodon retroversus) из ранней перми штата Техас, США. Американский музей естествознания, Нью-Йорк.
Ophiacodon hilli (Romer, 1925) Romer et Price, 1940 — вид, известный из ранней перми (сакмарский / артинский ярус, приуральский отдел) штата Канзас, США. Имеется неполный скелет.
Ophiacodon major Romer et Price, 1940 — вид, известный из ранней перми (сакмарский - кунгурский ярусы, приуральский отдел) штатов Оклахома и Техас, США. Найден фрагментарный материал (позвонки и фрагменты нижней челюсти). Будучи наиболее поздним, этот вид также является самым крупным представителем рода: его общая длина могла достигать 3,5-4 м и даже больше.
Существует также целый ряд остатков, определенных лишь до рода (Ophiacodon spp.), происходящий из ранней перми (ассельский - кунгурский ярусы, приуральский отдел) штатов Канзас, Колорадо, Нью-Мексико, Оклахома, Техас и Юта, США.

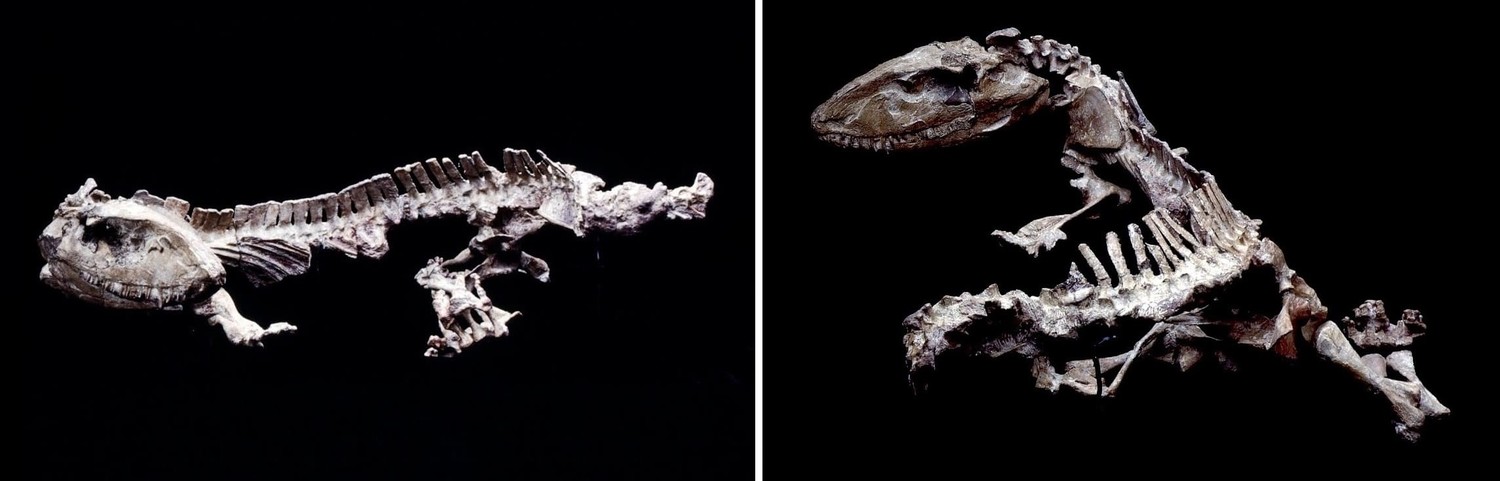
Скелет офиакодона (Ophiacodon sp.) из ранней перми севера штата Техас, США.
Ophiacodon beckiae Wilson, 1989 — некорректно описанный вид (nomen vanum), из позднего карбона (гжельский ярус) штата Канзас, США.
? Ophiacodon sp. Paton, 1974 — пеликозавр, описанный из ранней перми (предположительно ассельский ярус, приуральский отдел) Западной Англии, Великобритания. Найден плохо сохранившемуся фрагменту левой зубной кости. Изначально Мурчисон и Стрикленд (1840 г.) приняли этот образец за верхнечелюстную кость рыбы. Челюсть тонкая и очень низкая, постепенно расширяющаяся спереди назад. Сохранен симфизный отдел и полный зубной ряд, но большая часть вентральной части отсутствует. Длина фрагмента составляет 122 мм. В зубном ряду размещалось приблизительно 34 зуба. 1-й зуб в ряду, вероятно, был больше, чем 2-й и 3-й. 3-й зуб полностью сохранился, его длина составляет 11 мм. 4-я альвеола, вероятно, также вмещала большой зуб. Зубы с 5-го по 26-й примерно одинаковы по размерам, длиной 5 мм. Зубы с 27-го по 34-й намного меньше, уменьшаются в размерах спереди назад; длина наименьшего зуба — 2 мм. Зубы не сжаты с боков и не имеют режущих краев; зубы с 5-го по 34-й конические и колышковидные, с тупыми вершинами. Полностью сохранившийся увеличенный 3-й зуб немного загнут и заострен.

Зубы офиакодона (Ophiacodon sp.).
Структура челюсти и строение зубов английского Ophiacodon sp. типичны для офиакодонтов и отличаются от таковых у сфенакодонтов. Группа увеличенных зубов на переднем конце нижней челюсти характерна для пеликозавров. Зубы мельче и более многочисленны, чем у сфенакодонтов, а в зубном ряду есть много промежутков. Это также особенность, присущая офиакодонтам, и, вероятно, связанная с медленной сменой зубов. По структуре челюсть напоминает нижнюю челюсть пеликозавров рода Ophiacodon. До обнаружения этой находки представители рода, даже наиболее примитивные формы из карбона, были известны лишь в Северной Америке. Длина черепа европейского офиакодона могла равняться приблизительно 185 мм, что значительно меньше, чем у любой из известных американских форм. Однако, возможно, остатки принадлежат молодому животному. Количество зубов также меньше, чем у американских видов, хотя и превышает число, характерное для сфенакодонтов. Кроме меньшего размера и числа зубов, в остальном челюсть наиболее сходна с челюстью Ophiacodon uniformis, формы из асельского яруса формации Уичита. Слишком малое число отличительных признаков не позволяет отнести европейскую форму к отдельному или одному из описанных видов рода Ophiacodon.
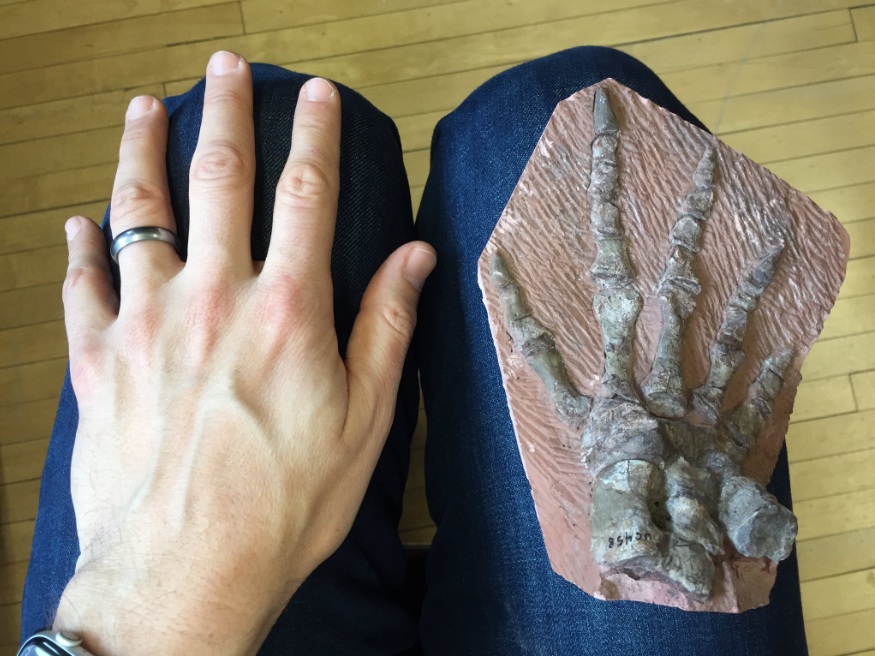
Скелет кисти офиакодона (Ophiacodon sp.).
Образ жизни
Наиболее вероятно, что офиакодоны жили на суше, хотя прежде их обычно считали полуводными. Водная среда обитания для этих животных впервые была предложена в 1907 г. Э. Кейзом, хотя позже он отказался от этой идеи. Анатомические особенности, которые могут указывать на то, что офиакодоны большую часть своего времени проводили в воде, включали уплощенные когтевые фаланги пальцев, приспособленные к загребанию воды, сильно вытянутые и узкие челюсти с многочисленными мелкими зубами, служившие адаптацией к питанию рыбой, а также слабо окостеневшие элементы запястья и предплюсны, встречающиеся у многих вторичноводных позвоночных. В 1940 г. А. Ромер и Л. Прайс дополнительно предположили, что более длинные задние конечности использовались для проталкивания тела сквозь водную толщу.

Офиакодон (Ophiacodon sp.) с добытой двоякодышащей рыбой. Автор — Р. Евсеев.
Некоторые из перечисленных признаком больше не считаются подтверждением полуводного образа жизни офиакодонов. Например, широкие когтевые фаланги пальцев встречаются у большинства ранних тетрапод, даже у тех, которые, как установлено, были почти исключительно наземными, а длинные задние конечности офиакодонов не могли служить им эффективным средством передвижения в воде, поскольку небольшие ступни образуют слишком маленькие гребные поверхности. Кроме того, у крупных особей слишком высокий череп, что не характерно для хищников-ихтиофагов. Анализ позвонков также указывает на то, что офиакодоны могут являться преимущественно наземными. Судя по всему, они питались самой разнообразной добычей — например, относительно небольшими амфибиями или дохлой рыбой.

Офиакодоны (Ophiacodon sp.) в естественном окружении. Автор — Ntvtiko.
Независимо от характера питания, офиакодоны являлись одними из крупнейших хищников своего времени; возможно, им приходилось опасаться лишь своих более крупных сородичей.


Офиакодон (Ophiacodon sp.) пытается не дать отнять сородичам двоякодышащую рыбу. Автор — Д. Гендерсон.
Ссылки
Офиакодон (Википедия) перейти
Офиакодонты (Википедия) перейти
Ophiacodon (Paleobiology Database) перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Иванов А. О., Черепанов Г. О. Ископаемые высшие позвоночные. — СПб.: Издательство Санкт-Петербургского университета, 2001
Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1 / Отв. ред. Ивахненко М. Ф. и Курочкин Е. Н. — М.: ГЕОС, 2008
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Павлова М. В. Палеозоология. Ч. 2. Позвоночные. — М.-Л.: Государственное издательство, 1929
Ромер А. Ш. Палеонтология позвоночных. — М. — Л.: Государственное научно-техническое издательство нефтяной и горно-топливной литературы, 1939
Самое древнее ископаемое яйцо. Природа 7, 1935
Татаринов Л.П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГЕОС, 2009
Paton R. L. Lower permian pelycosaurs from the English Midlands. Palaeontology 17 (3), 1974