Палеокасторы — Palaeocastor
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Суперотряд Эвархонтоглиресы (Euarchontoglires)
Грандотряд Глириобразные (Gliriformes)
Клада Глиресы (Glires)
Миротряд Симплицидентаты (Simplicidentata)
Отряд Грызуны (Rodentia)
Подотряд Боброобразные (Castorimorpha)
Надсемейство Кастороиды (Castoroidea)
Семейство Бобровые (Castoridae)
Подсемейство †Палеокасторины (Palaeocastorinae)
Род †Палеокасторы (Palaeocastor)
Вид †Palaeocastor nebrascensis
Вид †Palaeocastor wahlerti
Вид †Palaeocastor peninsulatus
Палеокасторы (Palaeocastor Leidy, 1869) — род подсемейства палеокасторин (Palaeocastorinae Martin , 1987), происходящий из раннего - позднего олигоцена Северной Америки и позднего олигоцена Азии. Установлены 3 вида.
Реконструкция внешнего облика палеокастора (Palaeocastor peninsulatus). Автор — Н. Тамура.
Синонимы: Castor Linnaeus, 1758 (частью); Steneofiber Geoffroy, 1833 (частью); Capacikala Macdonald, 1963 (частью).
История изучения
Голотип рассматриваемого рода был описан в 1856 г. Джозефом Лейди в качестве вида Steneofiber nebrascensis; таким образом, этот преимущественно североамериканский ископаемый бобр был вначале причислен к роду своих евразиатских сородичей. В 1869 г. Лейди в работе, описывающей окаменелости из Южной Дакоты и Небраски, дал более подробное описание вида и определил его в качестве типа нового рода, которому дал название Palaeocastor. Его образуют древнегреческие слова palaios — "древний" и castor — "бобр".
В 1905 г. Петерсон описал найденные остатки бобров в качестве 2 новых видов рода Steneofiber, но в дальнейшем они были отнесены к роду Palaeocastor. Так, вид Steneofiber barbouri чуть позже стал Palaeocastor barbouri, однако затем был отнесен к роду Pseudopalaeocastor. Аналогичным образом, таксон Steneofiber gradatus, выделенный Копом в 1879 г., временно побывал в составе рода Palaeocastor, но позже был реклассифицирован в Cpacikala. В последующие годы различными авторами выделялись многочисленные виды данного рода, в том числе Palaeocastor fossor, Palaeocastor nebrascensis, Palaeocastor magnus, Palaeocastor milleri, Palaeocastor peninsulatus и Palaeocastor simplicidens, часть из которых была в итоге выделена в отдельные роды (Pseudopalaeocastor, Fossorcastor), а другие определены как синонимичные.
В 1987 г. Ларри Мартин выделил в семействе бобровых новое подсемейство Palaeocastorinae, к которому отнес 4 описанных ранее рода ископаемых бобров (Palaeocastor, Capatanka, Capacikala и Euhapsis) и 2 новых. Рода Palaeocastor и Capatanka в классификации Мартина образуют трибу Palaeocastorini. В качестве общих признаков для этого подсемейства, представители которого были приспособлены к рытью нор и жизни в них, Мартин выделил уплощенные резцы и короткий, круглый в сечении хвост. По мнению этого автора, норные бобры были тупиковой боковой ветвью в своем семействе, и ни одна из последующих его ветвей от них не происходит. Видовая и даже родовая классификация внутри подсемейства Palaeocastorinae с тех пор оставалась проблематичной, и в 2010 г. Клара Стефен указывала, что эту классификацию трудно убедительно обосновать такими характеристиками, как форма зубов и форма и размер черепа.
Находки ископаемых остатков палеокасторин известны главным образом из западной части Северной Америки, происходя в основном из американских штатов Небраска, Вайоминг, Южная Дакота и Монтана. Самые ранние находки включают несколько частичных скелетов особей разного возраста из формации Форт-Логан в Монтане, которые датируются возрастом 29-27 млн. лет (поздний олигоцен). Столь же древними являются находки в лептаухениевом костеносном слое формации Шарпс, обнажения на берегу реки Уайт-Ривер близ деревни Вундед-Ни в Южной Дакоте, и откуда берет начало типовой материал рода Palaeocastor — почти полный череп и нескольких других черепных фрагментов. Кроме того, частичный череп, был также обнаружен в вышележащем слое формации Монро-Крик.
Выдающееся значение для исследования палеокасторин имеет формация Харрисон, отложения которой перекрывают формацию Монро-Крик и распространены на обширных пустынных территориях западной Небраски и соседнего восточного Вайоминга. Они представляют собой массивные, серо-коричневые и мелкозернистые песчаники, частично образовавшиеся в бывшем русле реки, кроме лесса, принесенного ветром. Эти слои относятся к раннему миоцену, с помощью радиометрических измерений по слоях вулканического пепла было выяснено, что они отлагались в период от 22,9 до 21,3 млн. лет (начало раннего миоцна). В осадочных породах с включением лесса были найдены многочисленные остатки Fossorcastor, включая полные скелеты, а также тысячи их окаменелых нор. История открытия и изучения последних представляет собой особый интерес.
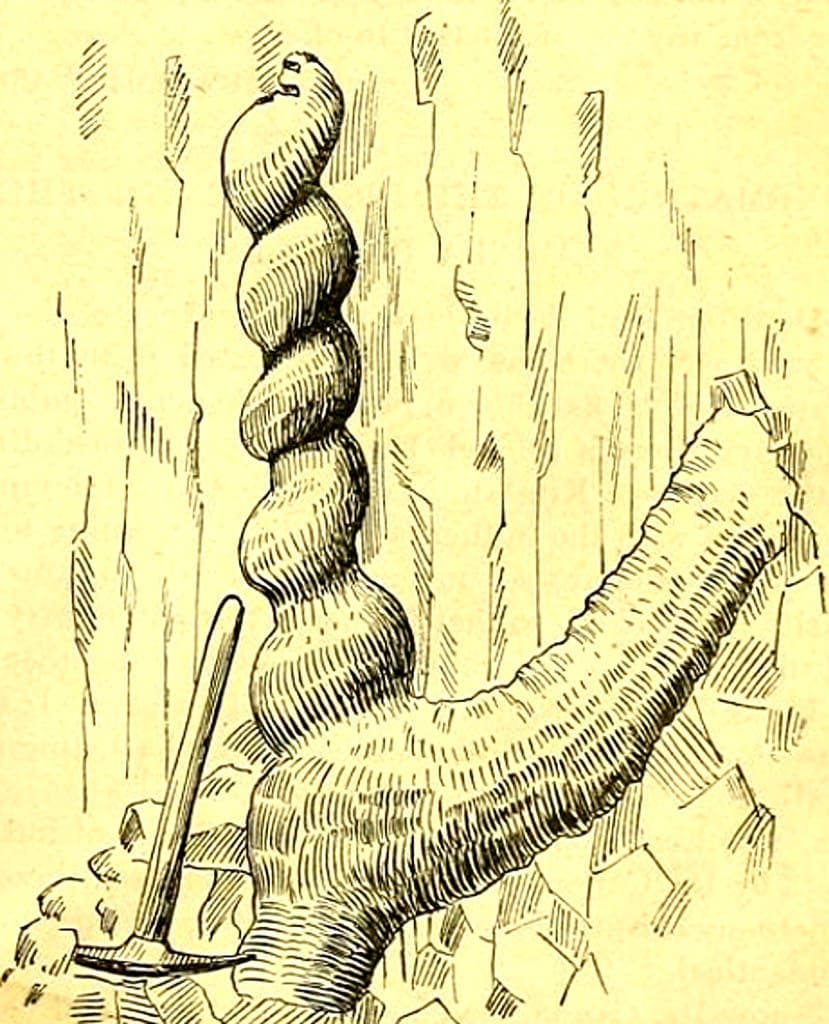
"Дьявольские штопоры" (помечены белыми стрелками), которые стали заметны благодаря эрозии раннемиоценовых отложений на склоне холма близ реки Найобрэра в штате Небраска, США.
В середине 1800-х гг. ковбои и первые владельцы ранчо в округе Сиу на северо-западе Небраски начали находить весьма странные образования из песчаника, торчащие из эродированных склонов прерий. Похоже, это были камни, но в отличие от других камней, виденных скотоводами раньше, они представляли собой трубки высотой с человека и толщиной с его руку, которые закручивались в идеальные спирали, как макароны фузилли. Снизу эти спирали заканчивались отходящим в сторону расширенным "корнем". Согласно раннему отчету, эти винтовые структуры были в некоторых местах настолько многочисленными, что, казалось, образовывали "настоящий лес" вдоль прерий. На территории площадью примерно 500 кв. миль их находилось около миллиона. Возникало ощущение, что какой-то великан с огромной силой ввинчивал в землю штопор, а затем вынимал его и заполнял получившиеся полости чем-то похожим на цементный раствор. Озадаченные фермеры стали называть эти странные сооружения "дьявольскими штопорами" (англ. devil’s corkscrews), и это название прижилось.
В 1891 г., когда американский геолог и палеонтолог Эрвин Хинкли Барбур проводил экспедицию в бедлендах в районе реки Уайт-Ривер, владелец ранчо Джеймс Кук обратили его внимание на "дьявольские штопоры" на своем участке, расположенном на берегу реки Найобрэра. Барбур выкопал несколько спиралей из земли и обнаружил, что они имеют наружные стенки из белого волокнистого материала, а внутри заполнены кальцинированными корнями, песком и илом. Конечно, как многие ученые своего времени, он являлся материалистом и не верил, что происхождение спиралей может иметь какое-то мистическое объяснение. Основываясь на том, что указанная местность в то время считалась дном огромного пересохшего древнего озера, ученый решил, что открыл новый, до этого неизвестный род ископаемых пресноводных губок, который он в небольшой статье в журнале «Science» за 1892 г. окрестил Daemonelix, переведя с английского на латынь народное название. Также Барбур отмечал, что по крайней мере в одной губке запутались кости вымершего грызуна.
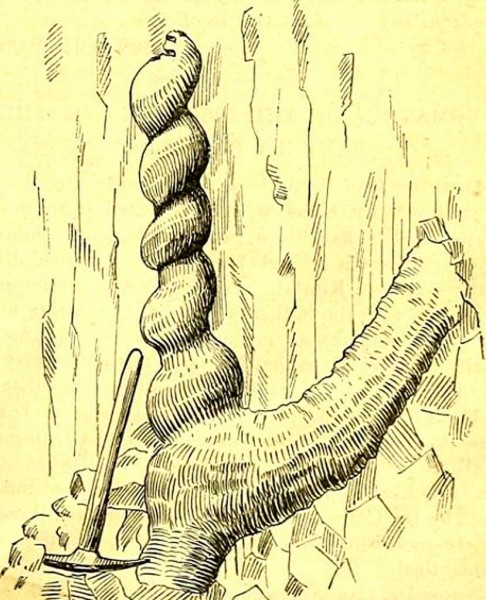
Рисунок из статьи Э.Х. Барбура в журнале «Science» за 1892 г., изображающий Daemonelix.
В 1893 г. американский палеонтолог позвоночных Эдвард Дринкер Коп отверг интерпретацию винтообразных окаменелостей Барбуром, отметив, что «наиболее вероятным объяснением этих объектов является то, что они являются слепками норок какого-то крупного грызуна». В том же году австрийский палеонтолог Теодор Фукс, специалист по ихнофоссилиям, независимо пришел к такому же выводу. Он отмечал, что «таким образом, мы вправе рассматривать эти странные окаменелости как не более чем подземные жилища миоценовых грызунов, вероятно, связанных с Geomys» [восточные гоферы (Geomys) — современный род североамериканских роющих грызунов семейства гоферовых (Geomyidae)]. Мелкие выпуклости, замеченные на некоторых спиралях, интерпретировались как следы когтей, оставленными зверьками во время рятья.
Однако профессор Барбур категорически отверг подобную идею и опубликовал критический ответ на статью Фукса в 1894 г. Предполагая, что вмещающие Daemonelix породы формации Харрисон являются озерными отложениями, Барбур прокомментировал: «Суслику доктора Фукса остается только зарыться и построить гнездо из сухого сена на глубине ста или двухсот саженей миоценовой воды». (Фукс сомневался, что окружающие скалы были озерными отложениями и интерпретировал остатки растений, найденные Барбуром, как сено, заготовленное строителями нор.)
"Дьявольские штопоры", обнаруженные в конце XIX в. на территории национального заповедника Агат-Фоссил-Бедс в штате Небраска, США.
Когда стало ясно, что местность, где находили окаменелости Daemonelix, была в раннем миоцене не озером, а полузасушливой степью, Барбур не сдался и в 1896 г. на заседании Американского геологического общества высказал предположение, что его находки являются не гигантскими губками, а корнеплодами неизвестных наземных растений, которые подверглись кальцинированию и поэтому смогли сохраниться. В подтверждение первооткрыватель представил впечатляющие образцы идеальных по форме "дьявольских штопоров" и заявил, что животные попросту не способны прорыть туннели столь совершенной формы, соблюдая такую симметрию. Барбур также заявлял, что открытие окаменелостей грызуна не является доказательством происхождения Daemonelix, поскольку также были найдены кости «млекопитающего размером с мышь». Правда, вскоре в ученом мире узнали, что для демонстрации Барбур отобрал самые лучшие образцы окаменелых спиралей, наиболее вписывающиеся в его "растительную теорию". Хотя существовали и другие "штопоры" — гораздо менее правильных форм. И все же возразить ученому с мировым именем тогда никто не посмел, а потому его теория главенствовала еще 8 лет.
В 1904 г. американский палеонтолог Олаф Петерсон собирал образцы Daemonelix для Музея естественной истории Карнеги в Питтсбурге, Пенсильвания. Он исследовал окаменелости и обнаружил в нижнем ответвлении одного из "штопоров" почти полный скелет ископаемого бобра. И тут все встало на свои места. Подобно старым канализационным трубам, норы были выстланы корнями (Барбур был прав относительно растительной ткани). Благодаря высокому содержанию кремния в грунтовых водах, корни растений превратились в белесую стеклянистую массу. Таким образом, Петерсон поддержал интерпретации Копа и Фукса. Со временем большинство исследователей, включая бывшего ученика Барбура и его преемника в Государственном музее К. Бертрана Шульца, признали спирали окаменелыми норами грызунов. Это самые ранние ископаемые норы млекопитающих, которые были распознаны и описаны.
Таким образом, винтообразные структуры оказались окаменелыми норами небольших древних бобров, ведущих исключительно наземный образ жизни. У них не было таких мощных хвостов, как у современных сородичей, однако имелись сильные передние конечности и челюсти, удобные для рытья. Небольшие бороздки на стенках нор (в негативе — выпуклости на поверхности) представляют собой следы резцов и когтей, которыми животные скребли мягкий грунт. При этом среди них были как "правши", так и "левши", в связи с чем ходы заворачивались в разные стороны. Нарытая земля затем выталкивалась на поверхность. Готовые норы заполнялись песком и илом во время паводков после сильных дождей, а они впоследствии сцементировались и образовали спирали, более плотные, чем окружающая порода. Из некоторых жилищ их обитатели так и не успели спастись и были погребены в наносах; судя по всему, обычно это происходило во время летней спячки.

Разнообразие нор палеокасторин. На дне гнездовой камеры одной из них заметен скелет фоссорокастора (Fossorcastor fossor).
Но, как оказалось, совершенное учеными открытие было не совсем таковым. Индейцы лакота, населявшие эти места, давным-давно знали о существовании и "дедушек-бобров", и их спиралевидных нор. Местность, где обнаружились "штопоры", они называли Плохими Землями. Время от времени натыкаясь здесь на скелеты динозавров, индейцы считали, что те до сих пор представляют собой опасность. Ведь подземные монстры в какой-то момент могли снова ожить! Особенно люди боялись Унктейхилы — огромного водного змееподобного чудища, с которым постоянно сражался Вакинйан (Громовое Существо). Чтобы древние страшилища больше не выходили на поверхность, Громовое Существо бросило клич: «Есть ли смельчаки, готовые принести себя в жертву ради спасения мира?». На зов откликнулись бобры. Они согласились добровольно обратиться в камень, чтобы не допустить на поверхность скрывающееся в недрах земли зло. С тех пор индейцы почитают древних бобров, а их окаменевшие спиралевидные норы называют "бобровыми домиками". Кстати, "дедушки-бобры" способны защитить человека не только от чудовищ, но и от вредных заклинаний, а потому амулеты из бобровых зубов имели большую ценность.
Окаменелые норы палеокасторин удалось обнаружить и в других местах, но подавляющее большинство из них приурочено именно к раннемиоценовым отложениям формации Харрисон. В наши дни "штопоры дьявола" — одна из достопримечательностей расположенного в Небраске национального заповедника Агат-Фоссил-Бедс. Прогуливаясь по бедлендам, туристы и сейчас могут увидеть выглядывающие из осыпавшихся склонов причудливые спирали. А в местном музее хранятся и окаменелые кости их древних строителей.
Состав
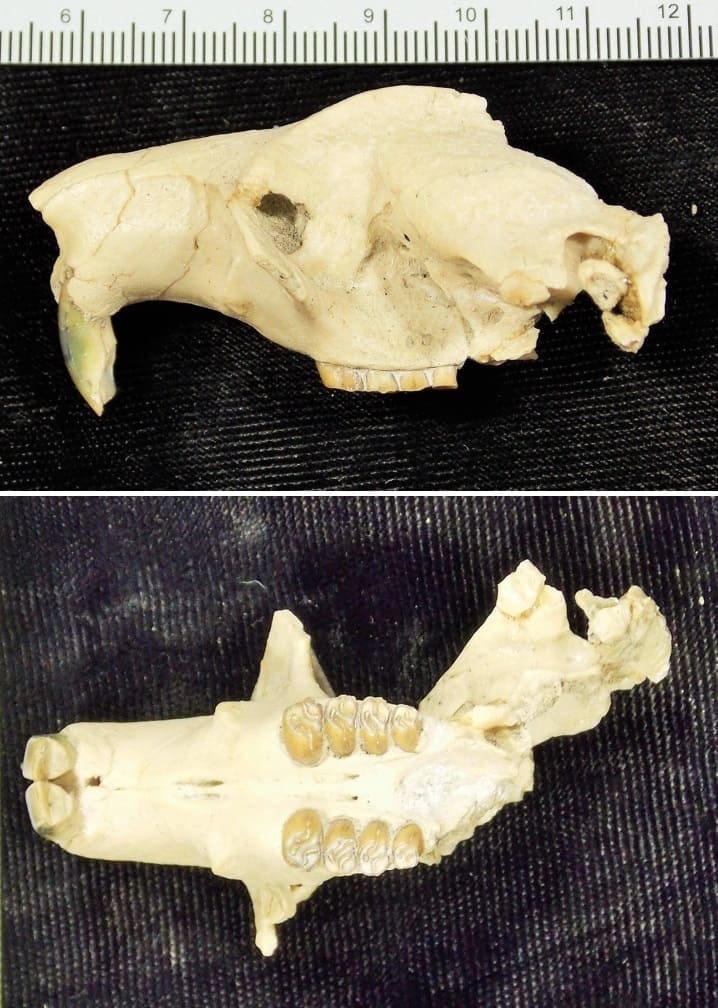
Palaeocastor nebrascensis (Leidy, 1856) Leidy, 1869 — типовой вид, известный из раннего - позднего олигоцена (уайтский - харрисонский века) США (штаты Вайоминг, Южная Дакота) и позднего олигоцена (хаттский ярус) Казахстана (Актюбинская область). Синонимы: Steneofiber nebrascensis Leidy, 1856 (тип); Steneofiber sciuroides Matthew, 1907; Capacikala sciuroides Macdonald, 1963.
Череп с нижней челюстью и элементы посткраниального скелета палеокастора (Palaeocastor nebrascensis) из олигоцена штата Южная Дакота, США. Длина образца — около 4 см.
Palaeocastor wahlerti Korth, 2001 — вид, известный из раннего олигоцена (уайтский век) США (штат Южная Дакота).
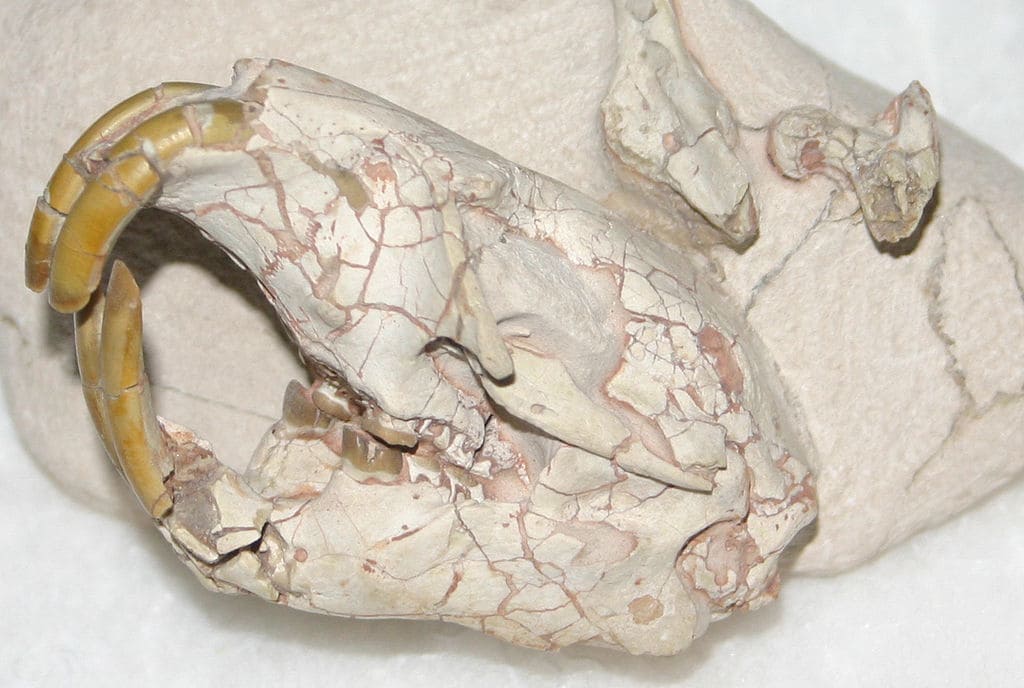
Palaeocastor peninsulatus (Cope, 1881) Stirton, 1935 — вид, известный из позднего олигоцена (герингский век) США (штаты Орегон, Южная Дакота). Синонимы: Castor peninsulatus Cope, 1881; Steneofiber peninsulatus Hay, 1902.
Череп с нижней челюстью палеокастора (Palaeocastor peninsulatus).
Остатки, определенные лишь до рода и обозначенные Palaeocastor spp., известны из позднего олигоцена (герингский - харрисонский века) США (штаты Небраска, Орегон, Южная Дакота) и позднего олигоцена (хаттский ярус) Казахстана.
Строение и функции
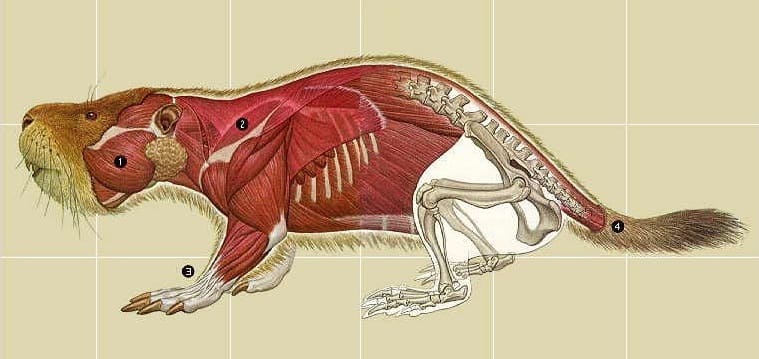
Палеокасторины выглядели как нечто среднее между ныне живущими бобрами и сусликами. Для них характерны крупные резцы, маленькие глаза и ушные раковины, мощные передние конечности и укороченный хвост. Морфология этих грызунов свидетельствует о возрастающей со временем приспособленности к рытью нор и жизни в них.
Реконструкция внешнего облика Palaeocastor. Расстояние между линиями сетки — 5 см.
Представители рода Palaeocastor имели средние для грызунов размеры: общая длина их тела достигала около 40 см , высота в наивысшей точке — порядка 12 см, а масса варьировала в пределах 0,8-1,2 кг. У более крупного Fossorcastor fossor, ранее включаемого в рассматриваемый род, масса тела могла достигать 4 кг.
Длина черепа составляетот 5,9 до 8,2 см, а его ширина в скуловых дугах — от 4,3 до 6,6 см. Череп Fossorcastor fossor достигает 9,2 см в длину и 8,8 см в ширину. Череп крупный по отношению к туловищу, но при этом сравнительно короткий, широкий и низкий, что характерно и для ныне живущих роющих грызунов — в частности, землекоповых (Bathyergidae). Морда сравнительно длиннее и уже, чем у современных бобров. Носовые и верхнечелюстные кости, образующие переднюю часть головы, несут небольшие утолщения, но не настолько развитые, как у ныне живущих под землей грызунов. Центральная часть предчелюстной кости, поддерживающая в основном крупные резцы, также существенно усилена. Более низкое расположение на черепе подглазничного отверстия является примитивным по сравнению с современными бобрами и даже с менее древними представителями подсемейства Palaeocastorinae.
Надбровные дуги, сходясь, образуют умеренно развитый сагиттальный гребень в месте наименьшего расстояния между глазницами или выше него. Лобная кость широкая и очень короткая; эта особенность выражена тем сильнее, чем в большей степени животные вели роющий образ жизни. На задней стороне теменных костей имеются два поперечных гребня, служащих дополнительными точками прикрепления мышц. В отличие от современных бобров и близких к ним форм, затылочная кость занимает всю заднюю часть черепа и занимает в основном вертикальное положение. Слуховые капсулы высокие и закругленные. У живущих под землей фоссоркасторов череп обычно имеет очень массивную структуру с мощными расширенными скуловыми дугами и высоким сагиттальным гребнем.
Неполный череп палеокастора (Palaeocastor peninsulatus).
Нижняя челюсть короткая и мощная, длиной от 5,9 до 6,4 см. Ее массивный симфиз доходит примерно до половины хорошо выраженной диастемы между резцами и щечными зубами. На нижней стороне челюсти расположен костный нарост, к которому прикреплялась двубрюшная мышца, что также характерно для современных бобров. Под р4 находится большое подбородочное отверстие. Приспособление к подземному образу жизни в основном выражается в увеличении глубины и ширины нижнечелюстного тела. В целом нижняя челюсть более примитивная, чем у современных бобров, со сдвинутой вперед восходящей ветвью и не столь сильно, как у более поздних родов, смещенным к средней оси суставным мыщелоком.
Подобно современным бобрам, зубная формула составляет I1/i1, C0/c0, P1/p1, M3/m3 = 20. Все зубы с очень высокой коронкой - гипсодонтные. Как и у современных бобров, резцы имеют долотообразную форму, их длина достигает 2,5 см в нижней челюсти и немного меньше в верхней. Эти зубы расположены в очень прочных трубчатых альвеолах и наклонно выступают вперед, особенно в верхней челюсти. В целом они слегка изогнуты и уплощены спереди, верхний край в основном имеет сильные скосы. Резцы быстро росли, чтобы противодействовать износу, возникающему в результате рытья ими почвы.
За резцами в верхней челюсти находится большая диастема, длина которой может достигать 3 см, в нижней челюсти диастема вдвое меньше. Последний предкоренной самый большой в ряду щечных зубов, длиной 4-4,5 мм и с прямоугольным контуром жевательной поверхности. Коренные зубы постепенно уменьшаются кзади и принимают квадратную форму, за исключением самого последнего, который имеет круглые или треугольные очертания. Длина всего ряда щечных зубов составляет 1,2-1,6 см. Рисунок их жевательной поверхности состоит из нескольких углублений, которые образованы из дентина и окружены эмалью. Форма и количество этих углублений сильно варьирует в зависимости от степени износа зубов.
Посткраниальный скелет палеокасторин хорошо известен. Позвоночник состоит из 7 шейных, 13 грудных, 6 поясничных, 5 крестцовых и, вероятно, 18 хвостовых позвонков. Особые отличия от современных бобров проявляются в хвостовом отделе позвоночника. Тела позвонков имеют округлую форму и практически не уменьшаются в размерах на всем протяжении хвоста. Поперечные отростки отчетливо выражены, проксимальные позвонки, возможно, имели шевронные отростки на нижней стороне, о чем свидетельствуют сохранившиеся там костные выпуклости.
Передняя конечность приспособлена к роющей деятельности. Ключица хорошо развита. Плечевая кость очень короткая и компактная, от 3,5 до 4,6 см в длину. Она имеет широкий нижний и узкий верхний суставный конец, снабженный большой суставной головкой. Локтевая кость составляет от 4,1 до 5,6 см в длину, вверху она довольно широкая, а к низу становится тоньше. В целом она уплощена с боков, длина верхнего суставного отростка составляет около 1 см. Лучевая кость немного легче локтевой, с круглым стержнем и большой головкой. Обе кости предплечья не сращены друг с другом.
Бедренная кость длиной от 4,8 до 5,4 см. Она также относительно стройная, ее стержень не настолько уплощен, как у современных бобров. Суставная головка расположена на отчетливой, хотя и короткой, шейке. Кости голени еще не демонстрируют заметного удлинения, как у современных бобров, и имеют несколько более короткую или ту же длину, что и верхний отдел конечности. Большая берцовая кость не сращена с малой, ее стержень округлый в поперечном сечении и имеет небольшой поворот по продольной оси. Кисть и стопа пятипалые. Пальцы крепкие, с короткими и широкими фалангами. Фаланги пальцев кисти по размерам превосходят таковые пальцев стопы.
В целом передние лапы палеокасторин более сильные, широкие и уплощенные, чем задние, а хвост укороченный и явно округлый в поперечном сечении (для сравнения, у современных бобров уплощены и передние, и задние лапы, что является адаптацией к водному образу жизни, а хвост широкий и расплющенный, напоминающий по форме лопасть весла).
Степень приспособленности к норному образу жизни у разных видов рода была разной — в частности, Palaeocastor peninsulatus был менее специализирован, чем более поздний Fossorcastor fossor, сохранив широкий зрительный канал, что свидетельствует о том, что он больше времени проводил на свету. Кроме того, у разных видов наблюдаются черты, указывающие на разный основной способ рытья, и если Fossorcastor fossor использовал для этого в основном резцы, то Palaeocastor peninsulatus свои норы, по-видимому, рыл преимущественно лапами.
Образ жизни
Некоторые палеокасторины устраивали норы необычной спиральной формы, часто намотанные вокруг вертикальной оси. В полностью сохранившихся норах количество витков может составлять от 6 до 12. Расположенный на поверхности единственный вход ведет в коридор, который составляет от 10 до 15 см в диаметре и проникает на глубину от 210 до 275 см со средним углом наклона от 25 до 30 °. Спиральный коридор заканчивается отходящей в сторону гнездовой камерой — горизонтальной или слегка наклонной (от 2 ° до 37 °), с самой низкой точкой у входа; в длину она может вытягиваться до 450 см при диаметре от 15 до 22 см. Это жилое помещение, именно здесь обычно находят скелеты роющих бобров и их детенышей. Некоторые, большей частью относительно пологие камеры имеют более мелкие карманы по бокам, которые могли служить кладовыми или уборными. Часть из нор содержат гнездовые камеры с большим наклоном (около 45 °), которые, возможно, защищали спящих грызунов от наводнения, что может указывать на наличие у них летней спячки (эстивации).
Норы роющих бобров были построены посредством поочередных ударов право- и леворасположенных резцов. Количество право- и левосторонних спиралей примерно одинаково. Должно быть, грызун фиксировал свои задние лапы на оси спирали и буквально ввинчивался в землю. Входы в норы были отмечены высокими холмиками из выкопанного грунта. Вероятно, бобры зачерпывали почву передними лапами и пробрасывали под брюхом назад. Время от времени грызуны, должно быть, использовал свою уплощенную голову, чтобы вытолкнуть нарытую землю из норы. Норные ходы небольшого Pseudopalaeocastor barbouri и относительно крупного Fossorcastor fossor можно отличить по диаметру спирали и ширине следов рытья.
Колония палеокасторов (Palaeocastor sp.). Автор — Р. Хорсфалл.
Примечательно, что норы луговых собачек (Cynomys) — современных представителей семейства беличьих (Sciuridae), во множестве населяющих североамериканские Великие равнины, включая Небраску, — за исключением спиралевидной формы, обладают с норами палеокасторин рядом общих черт. Эти грызуны, которые достигают размеров кролика, живут в сложном лабиринте подземных туннелей, часто большими колониями в сотни или даже тысячи особей, называемыми городками. Внутри этих защитных жилищ находится множество камер, которые используются для различных целей: хранения пищи, воспитания потомства, а также в качестве туалетов. Небольшое расстояние между норами, найденными в одном и том же месте, и их значительное количество свидетельствуют о том, что палеокасторины тоже вели общественный образ жизни. Подобно луговым собачкам, они могли устраивать обширные сети городков, занимающие целые акры. Обнаруженные вместе остатки взрослых и молодых особей подтверждают, что эти бобры выводили внутри подземных камер свой приплод. Возможно, некоторые зверьки стояли на страже на близлежащих холмах, чтобы предупреждать об опасности других членов колонии громким свистом.

Семейство палеокасторов (Palaeocastor sp.) в их спиралевидной норе. Самка присматривает за детенышами в гнездовой камере, в то время как самец следит за входом в нору. Автор — Б. Николлс.
Устройство спиралевидных нор требовало от палеокасторин бо́льших усилий, чем рытье прямых наклонных ходов такой же длины. В числе причин, приведших роющих бобров к такой форме, называется их территориальная скученность: в условиях больших колоний прямые норы должны были часто пересекаться. Согласно другому предположению, необычная форма нор была ответом на жаркий и сухой климат той эпохи. С этой точки зрения, ее спиралевидная конструкция могла являться своеобразной системой кондиционирования воздуха для сохранения оптимального режима температуры и влажности внутри гнездовой камеры, что было особенно важно для успешного выведения потомства. В пользу последней гипотезы также свидетельствуют спиралевидные норы ныне живущего в Австралии и на Новой Гвинее аргусова варана (Varanus panoptes), который использует их исключительно для гнездования.

Палеокасторы (Palaeocastor sp.), занятые рытьем нор. У одного из входов расположился зодиолест (Zodiolestes daimonelixensis) — хищник из семейства куньих, который мог охотиться на этих грызунов. Автор — Х. Ласерда.
Известно, что у роющих бобров имелись враги. Так, в одной из их окаменелых нор был найден свернутый калачиком скелет позднеолигоценового куньего зодиолеста (Zodiolestes daimonelixensis). Вполне вероятно, что этот хищный зверек отправился на поиски пищи в нору, вырытую бобрами, а затем оказался там в ловушке. Подобные ситуации в настоящее время происходят, например, с чернохвостыми луговыми собачками (Cynomys ludovicianus) и охотящимися на них черноногими хорьками (Mustela nigripes). Преследуемый на поверхности, палеокасторин может попытаться спастись, нырнув головой вперед в свою нору. Вершины нор открываются расширенными входами, которые позволили бы убегающему бобру развернуться, а затем высунуть голову над насыпью или отступить в нору, только немного более широкую, чем его тело, и встретить хищника сильными челюстями с большими резцами. Дополнительные выходы из гнездовой камеры отсутствуют, также подтверждая, что хозяева нор предпочитали не спасаться бегством от хищников (которые из-за небольшого диаметра хода не могли быть намного крупней их самих), а занимали оборону у входа в жилье.

Зодиолест (Zodiolestes daimonelixensis) почти настиг палеокастора (Palaeocastor sp.) в гнездовой камере его норы. Автор — Х. Ласерда.
Эволюция
В конце палеогена климат Северного полушария становится более прохладным и засушливым. Господствующим типом местности стали семиаридные степи, что заставило многих грызунов адаптироваться к норному виду жизни. Своя норная ветвь, основу рациона которой составляли ненасыщенные кремнием травы, выделилась и среди Castoridae. Сегодня это семейство насчитывает лишь 2 вида в пределах рода Castor. Однако оно имеет богатую эволюционную историю, восходящую к позднему эоцену, и поэтому включает множество родов в нескольких подсемействах. Современные бобры относятся к подсемейству Castorinae, представленное также более древними представителями их филогенетической линии. Castorinae вместе с сестринской группой Castoroidinae, в которую входят такие известные роды, как Castoroides и Trogontherium, объединяет полуводные формы. Род Palaeocastor, с другой стороны, относится к подсемейству Palaeocastorinae, которое существовало только в олигоцене и миоцене. В числе его ближайших родственников — Fossorcastor и Pseudopalaeocastor. Подсемейство Agnotocastorinae, в свою очередь, образует сестринскую группу по отношению к Palaeocastorinae. Представители обоих подсемейств приспособились к наземному образу жизни в открытых ландшафтах и иногда специализировались на рытье.
Согласно филогенетическим исследованиям, как полуводная группа (Castorinae и Castoroidinae), так и наземно-подземная группа (Palaeocastorinae и Agnotocastorinaee) образуют монофилетическую единицу, причем первая, согласно молекулярным данным, существует не менее 19,7 млн. лет. Поскольку нынешние бобры создают каналы и запруды по берегам рек, как и Steneofiber из раннего миоцена Европы и Nothodipoides из среднего миоцена Северной Америки, которые считаются полуводными, а также выкапывали норы, можно предположить, что общий предок обеих групп уже был приспособлен для рытья. Бобры, которые в основном рыли норы и населяли открытые ландшафты, скорее всего, вымерли в течение миоцена, конкурируя с другими млекопитающими, ведущими сходный образ жизни, такими как грызуны из семейства Mylagaulidae. Характерное грызение деревьев, по-видимому, развилось только дважды в пределах полуводной группы, у Castor и Dipoides (последний близок к Castoroides и Trogontherium). Возможно, подгрызание деревьев возникло в результате первоначального использования береговой кустарниковой растительности. Таким образом, поскольку эта особенность сформировалась в полуводной группе бобров, ее можно рассматривать как производную характеристику.
Ссылки
Palaeocastor (The Paleobiology Database)
перейти
Литература
Lomax D. R. Locked in time. Animal behavior unearthed in 50 extraordinary fossils. — New York: Columbia University Press, 2021