История изучения
Первое научное описание пальмовой циветы сделал в 1830 г. Джон Эдвард Грей, присвоив ей название Viverra binotata. Таксон был выделен по экземпляру, происходящему из области Ашанти в Гане. В 1832 г. этот же автор реклассифицировал животное в качестве Paradoxurus binotatus, одновременно описав другой вид — Paradoxurus hamiltonii. В 1843 г. он синонимизировал второй вид с первым и установил для него монотипический род Nandinia. В 1864 г. Грей объединил его с родами Paradoxorus, Paguma и уже невалидным Arctogale в трибу Paradoxurina в рамках семейства Viverridae. Томас в 1893 г. описал вид Nandinia gerrardi, Геллер в 1913 г. — Nandinia arborea, Кабрера и Ракстон в 1926 г. — Nandinia intensa. В дальнейшем они были признаны младшими синонимами вида Nandinia binotata и сейчас рассматриваются в качестве его подвидов.
В 1929 г. Реджинальд Иннес Покок выделил пальмовую цивету в собственное семейство Nandiniidae, таким образом дифференцировав ее от Viverridae. Автор обосновывал это тем, что это животное отличается от виверровых примитивными особенностями строения слухового барабана, сосцевидной части височной кости и пахучих желез. Однако большинство зоологов до конца ХХ в. продолжало относить пальмовую цивету к виверровым, в составе которых она выделялась в отдельное подсемейство Nandiniinae. Наличие у пальмовой циветы архаичных морфологических признаков позволило Херлуфу Винге в 1941 г. рассматривать ее в качестве единственного сохранившегося представителя семейства Amphictidae, содержавшего также такие ископаемые роды, как Amphictis, Miacis и Didymictis.
В настоящее время Nandinia binotata выделяется в монотипическое семейство Nandiniidae. Генетические исследования показали, что оно не является близкородственным к Viverridae, но образует независимую линию внутри клады Aeluroidea, представляя собой сестринскую группу по отношению к прочим ее представителям. Согласно молекулярным часам митохондриальной ДНК, отделение Nandiniidae от общего ствола произошло в самом конце эоцена, около 35 млн. лет назад. Это время ознаменовалось глобальным похололанием примерно на 5 °С и сокращением лесных массивов, вследствие чего многие группы млекопитающих вымерли и появились новые. Предки пальмовой циветы остались жить на деревьях в тропических лесах, тогда как другие кошкообразные начали осваивать более открытые ландшафты. Медленно эволюционируя в довольно стабильной среде, Nandiniidae не только сохранили много древних признаков, но и выработали ряд специализированных черт.
Строение и функции

Пальмовая цивета (Nandinia binotata).
Пальмовая цивета имеет небольшие для хищных размеры. В них наблюдается половой диморфизм: самцы обычно несколько крупнее самок. Длина головы и туловища составляет 39,8-62,5 (в среднем 48,6) см у самцов и 37-61 (в среднем 45,7) см у самок. Длина хвоста 43-76,2 (в среднем 53,8) см у самцов и 34-70,1 (в среднем 50,4) см у самок. Высота в плечах 8-13 см. Вес 1,3-3,2 (в среднем 2,1) кг у самцов и 1,2-2,7 (в среднем 1,94) кг у самок. По некоторым данным, вес самцов может достигать 5 кг. Длина ушной раковины 2,9-4,8 (в среднем 3,9) см у самцов и 2,7-4,3 (в среднем 3,7) см у самок. Длина стопы 7,6-10 (в среднем 8,8) см у самцов и 6,6-9,5 (в среднем 8,2) см у самок.

Портрет пальмовой циветы (Nandinia binotata).
Внешне животное несколько напоминает кошку. Голова широкая с умеренно заостренной мордой. Цвет ринариума варьирует от розовато-серого до темно-коричневого. Вибриссы хорошо развиты. Радужка глаз от желто-зеленой до оранжево-коричневой, зрачки вертикальные щелевидные. Ушные раковины широко расставленные, короткие, широкие у основания и с округлыми вершинами. Шея довольно длинная. Туловище вытянутое. Конечности короткие, задние примерно на 1/3 длиннее передних. Они очень сильные, но при этом обладают исключительной подвижностью в суставах. Кисть и стопа крупные, пятипалые. I-е пальцы на них меньше остальных пальцев и в некоторой мере противопоставляются им. Когти на всех пальцах острые, изогнутые, способные частично втягиваться. На подошвах и дистальных фалангах орошо развиты подушечки, облегчающие сцепление с субстратом. На кисти подушечка простирается до запястья, на стопе — до лодыжкки. На кисти и передней части стопы подушечка разделяется на отдельные крупные сегменты, а на задней части стопы покрыта сложным рисунком из многочисленных поперечно расположенных складок и пупырышков. Подушечки III-го и IV-го пальцев отделены от стопы широким пространством. Область между подошвенными и пальцевыми подушечками (за исключением I-х пальцев кисти и стопы) покрыта волосами. Передние конечности стопоходящие, задние полустопоходящие. Хвост на 10-20 % превышает по длине остальное тело; он довольно толстый, но при этом гибкий.
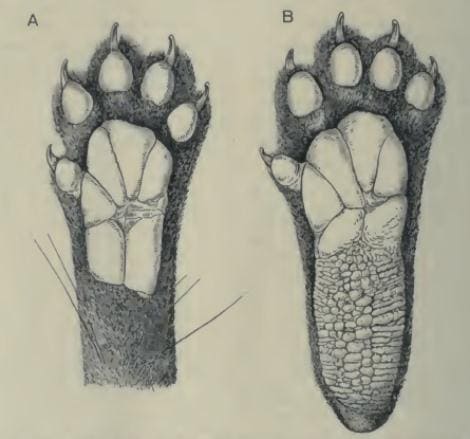
Прорисовки подошвенных поверхностей левых кисти (А) и стопы (В) пальмовой циветы (Nandinia binotata).
Шерстный покров короткий, густой и мягкий, но волосы имеют жесткие кончики. На хвосте волосы удлинены, что делает его пушистым. Окрас меха подвержен широкой географической и индивидуальной изменчивости. Сверху и по бокам головы, шеи, туловища и конечностей его общий фон тусклый, варьирующий от сероватого до темно-коричневого, с рыжеватым или каштановым оттенком. По бокам морды, над глазами и переносицей обычно имеются потемнения. За каждым ухом расположено темное пятно. Сверху и по бокам шеи выше линии ушей находятся 3-5 темных продольных полос, образованных в результате слияния пятен. Позади плеч на лопатках часто присутствуют 2 беловато-желтых пятна. На спине, по бокам туловища, снаружи плеч и бедер имеются нерегулярно расположенные и слабо выраженные темные пятнышки или "рябь", а на крестце и в основании хвоста — нечеткие темно-коричневые пятна. Брюшная сторона немного светлее спины и боков. Горло, как правило, сероватое. Брюхо и внутренние поверхности конечностей серые с желтоватым оттенком, без пятен. Хвост обычно несколько темнее остального тела, сходного цвета сверху и снизу, с изменчивой картиной из 9-15 размытых, обычно хуже выраженных снизу колец. Цвет пятен, полос и колец варьирует от темно-коричневого до черного; их количество, размеры и интенсивность окраса меняются от особи к особи. Альбинизм встречается крайне редко.
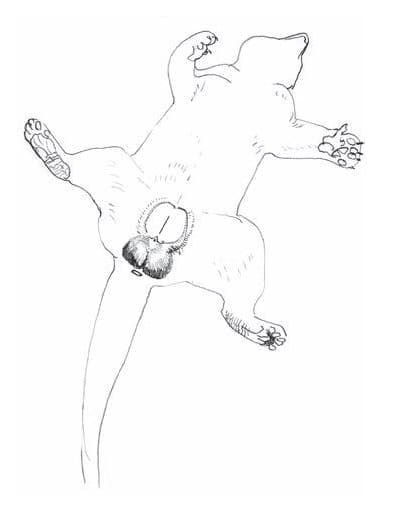
Рисунок брюшной стороны пальмовой циветы (Nandinia binotata).
У пальмовой циветы имеется много специфических пахучих желез. Одна из них расположена снизу на подбородке, другие — на подошвах лап. Между III-м и IV-м пальцами обеих пар конечностей в специальных карманах, под пятнами желтого меха, находятся железы, выделяющие секрет с фруктовым запахом. В складке безволосой кожи по средней линии в нижней части живота впереди препуция у самцов и вульвы у самок расположена непарная железа, вырабатывающая большое количество желтовато-коричневого секрета с довольно резким запахом. В открытом состоянии железистая зона составляет около 5,5 см в длину, 2-3 см в ширину и 2 см в глубину. По сравнению со сходной железой виверровых, она имеет меньшие размеры, более уплощенную форму, относительно простое строение и находится не в области промежности, между мочеполовым и анальным отверстиями, а перед гениталиями. Также у самок в толще кожи над молочными железами имеются пахучие железы, выделяющие в период лактации яркий оранжево-желтый секрет. Сосков 2 пары. У самцов имеется бакулюм.

Смонтированный скелет пальмовой циветы (Nandinia binotata).
Наибольшая длина черепа составляет 8,64-11,09 (в среднем 10,06) см у самцов и 8,44-10,32 (в среднем 9,47) см у самок. Наибольшая ширина черепа 5,32-6,27 (в среднем 5,72) см у самцов и 4,73-5,84 (в среднем 5,27) см у самок. У черепа умеренно вытянутый лицевой отдел и вытянутая мозговая капсула с хорошо выраженными стреловидным и затылочным гребнями. Скуловые дуги расставлены относительно широко. Костное небо треугольной формы. Слезная и скуловая кости не соприкасаются. Передний край глазницы образован лобной, верхнечелюстной и скуловой костями. Каменистая кость имеет строение, типичное для примитивных вымерших кошкообразных. Слуховой барабан также примитивен — не вздут и состоит из единственной камеры, без перегородки. У взрослых особей окостеневает только передняя часть барабана, тогда как расположенная каудально внутренняя барабанная кость навсегда остается хрящевой. Этот признак является плезиоморфным; он характерен для целого ряда ископаемых хищных, но почти не встречается у их современных представителей (исключение составляют лишь пальмовая цивета и некоторые особи мусангов (Paradoxurus spp.)). Внутреннее сонное отверстие расположено между основной клиновидной и внутренней барабанной костями. Околозатылочный отросток относительно толстый, длинный, остроконечный, направленный назад и не соприкасающийся со слуховым барабаном. Внешне он скорее напоминает аналогичный отросток собакообразных, а не прочих кошкообразных. Верхний заглазничный отросток крупный, нижний отросток небольшой; заглазничная дуга отсутствует.
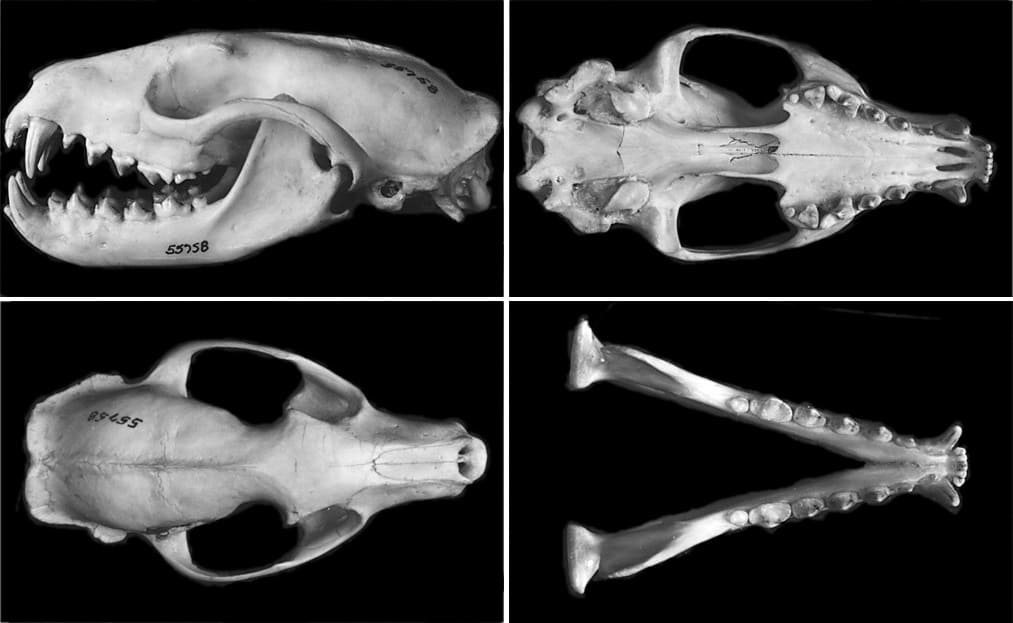
Череп с нижней челюстью пальмовой циветы (Nandinia binotata).
Зубная формула: I3/i3, C1/c1, P4/p4, M1-2/m1-2 = 36-40. Число щечных зубов может индивидуально врьировать. Из 121 исследованного экземпляра у 91 имелся полный возможный набор зубов. В то же время, у 10 экземпляров отсутствовали оба M2, у 4 — правый M2, у 2 — левый M2, у 1 — оба m2, у 1 — оба m2 и правый M2, у 1 — левый P1, у 1 — оба P1, у 3 — левый p1, у 3 — правый p1, у 2 — оба p1, у 1 — оба M2 и оба p1. У 2 экземпляров имелся дополнительный p1. Верхние резцы формируют поперечную линию. Как верхние, так и нижние клыки большие, прямые и относительно желобчатые (в большей степени, чем у генет (Genetta spp.)). Хищнические зубы имеют относительно примитивное строение. На P3 присутствуют лингвальный бугорок. M2/m2 сильно редуцированы и с округленными коронками, часто совсем отсутствуют. m2 с небольшим лингвальным бугорком.
Слепая кишка у пальмовой циветы отсутствует. Транспромонториальность в расположении внутренней сонной артерии выражена незначительно, что считается плезиоморфным для кошкообразных признаком. Средняя температура тела составляет 37,4 °С. Средний уровень основного обмена 5,57 куб. см кислорода на 1 г в 1 ч. Подобно другим хищным, хорошо развиты органы чувств: зрение, слух, осязание и, особенно, обоняние. Согласно наблюдениям, сделанным в неволе, по сообразительности пальмовая цивета превосходит большинство домашних собак и кошек.
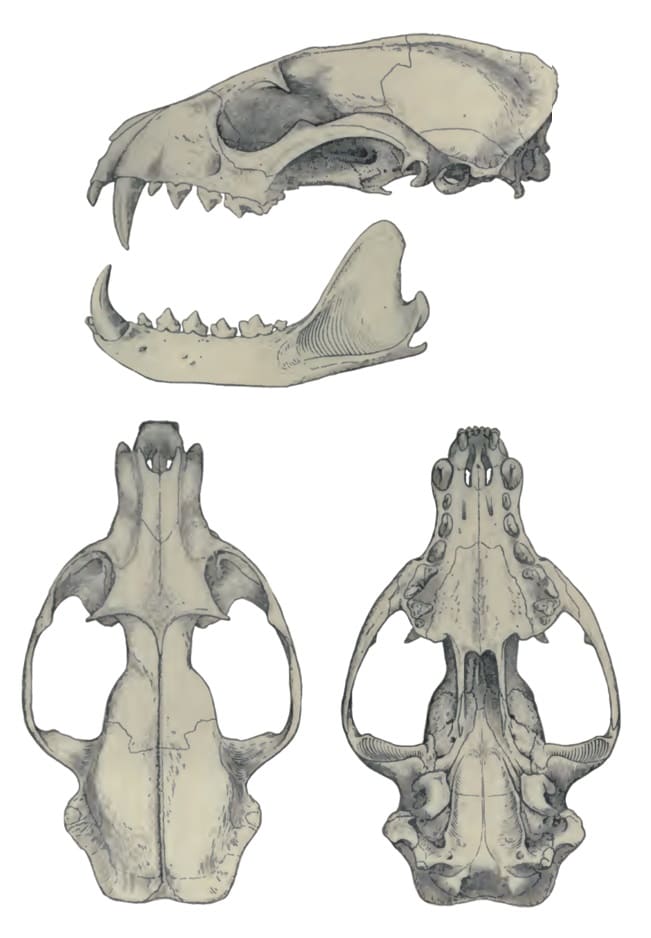
Прорисовки черепа с нижней челюстью пальмовой циветы (Nandinia binotata).
В кариотипе пальмовой циветы 38 хромосом и 66 хромосомных плеч. Расшифрованная митохондриальная ДНК состоит из 17 103 пар оснований и содержит 37 генов, типичных для генома млекопитающих. Из них 13 генов экспрессируют информационные РНК, которые кодируют белковые субъединицы, 22 — транспортную РНК и 2 — рибосомальную РНК.
Распространение
В настоящее время ареал пальмовой циветы охватывает южную половину Западной Африки и значительную часть Центральной и Восточной Африки. На востоке материка он теряет однородность и распадается на отдельные фрагменты. Пальмовая цивета встречается на юге Сенегала, в Гамбии, Гвинее-Бисау, Гвинее (кроме северо-востока), Сьерра-Леоне, Кот-д'Ивуар (кроме крайнего севера), Либерии, центре и юге Ганы, юге Того, юге Бенина, юге Нигерии, Камеруне (кроме севера), Экваториальной Гвинее (на о. Биоко в Гвинейском заливе, вероятно, уже исчезла), Габоне, Республике Конго, юге Центральноафриканской Республики, Демократической Республике Конго (кроме юго-востока), севере Анголы (южная граница около 9° ю. ш.), юге Южного Судана, Уганде (кроме северо-востока), Руанде, Бурунди, западе и юго-западе Кении, на северо-западе, северо-востоке, востоке, в центре и на юго-западе Танзании (включая остров Занзибар), крайнем северо-западе и северо-востоке Замбии, большей части Малави (кроме запада), севере и западе Мозамбика (южная граница около 20°45′ ю. ш.), крайнем востоке Зимбабве.
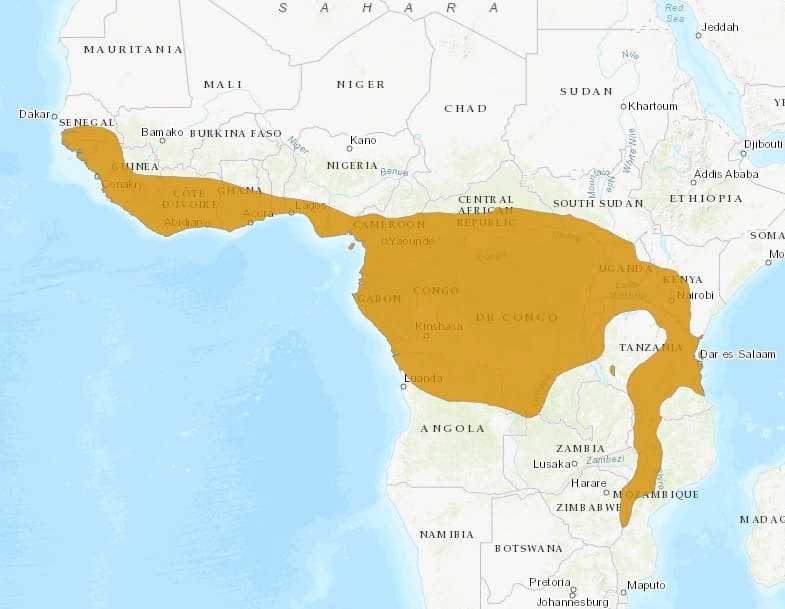
Современный ареал пальмовой циветы (Nandinia binotata).
Состав
На основе географических вариаций окраса шерсти условно выделяют 4 подвида пальмовой циветы.
Nandinia binotata binotata (Gray, 1830) — большая часть ареала, от Гамбии на восток и юг до Камеруна, Экваториальной Гвинеи (включая о. Биоко), Габона, Конго, юга Ценртальноафриканской Республики, Демократической Республики Конго и севера Анголы. Номинативная форма, внешний облик которой наиболее типичен для вида.

Пальмовая цивета (Nandinia binotata binotata) из Габона.
Nandinia binotata arborea (Heller, 1913) — юг Южного Судана, Уганда, юго-запад Кении и север Танзании (до течения реки Руфиджи; горы Килиманджаро, Меру и Восточный рифт). Темные полосы на шее и кольца на хвосте узкие.
Nandinia binotata gerrardi (Thomas, 1893) — юго-восток Танзании (от течения реки Руфиджи), Малави, крайний северо-запад Замбии (район Мвинилунга), восток Зимбабве, запад Мозамбика; предположительно, также остров Занзибар. Темные полосы на шее отсутствуют. Светлые пятна на лопатках слабо выраженные. Темные пятна на теле относительно мелкие и редко расположенные. Темные кольца на хвосте отчетливо выраженные, узкие и расположенные близко друг к другу.
Nandinia binotata intensa (Cabrera et Ruxton, 1926) — юг Демократической Республики Конго, север Анголы и крайний северо-восток Замбии (плато Ньика). Окрас верхней и нижней стороны тела сравнительно яркий, с рыжеватым оттенком. Светлые пятна на лопатках отчетливо выраженные. Темные пятна насыщенно черные.
Согласно имеющимся данным, достаточных оснований для выделения всех указанных форм не существует. Изучение более 100 шкурок, добытых в различных районах Африки, позволяет считать Nandinia binotata gerrardi и Nandinia binotata intensa младшими синонимами Nandinia binotata binotata. Четкая граница между Nandinia binotata gerrardi и Nandinia binotata arborea также отсутствует: у особей, добытых в горах Удзунгва (центральная часть Танзании), светлые пятна на лопатках хорошо выражены или совсем отсутствуют.