Паракамелюсы — Paracamelus
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Клада Скротиферы (Scrotifera)
Клада Феревунгуляты (Fereuungulata)
Клада Эвунгуляты (Euungulata)
Клада Параксонии (Paraxonia)
Отряд Парнопалые (Artiodactyla)
Подотряд Мозоленогие (Tylopoda)
Надсемейство Камелоиды (Cameloidea)
Семейство Верблюдовые (Camelidae)
Подсемейство Камелины (Camelinae)
Триба Камелини (Camelini)
Род †Паракамелюсы (Paracamelus)
Вид †Paracamelus aguirrei
Вид †Paracamelus longipes
Вид †Paracamelus praebactrianus
Вид †Paracamelus gigas
Вид †Paracamelus trofimovi
Вид †Paracamelus сhersonensis
Вид †Paracamelus alexejevi
Вид †Paracamelus alutensis
Паракамелюсы (Paracamelus Schlosser, 1903) — род подсемейства камелин (Camelinae Zittel, 1893), происходящий из среднего миоцена - позднего плиоцена Северной Америки, раннего плиоцена - среднего плейстоцена Азии, позднего плиоцена - раннего плейстоцена Европы и позднего миоцена Африки. Установлено около 8 видов.

Реконструкция внешнего облика паракамелюса (Paracamelus gigas). Автор — Р. Евсеев.
Синонимы: Camelus Linnaeus, 1758 (частью); Gigantocamelus Barbour et Schultz, 1939 (частью).
Строение и функции
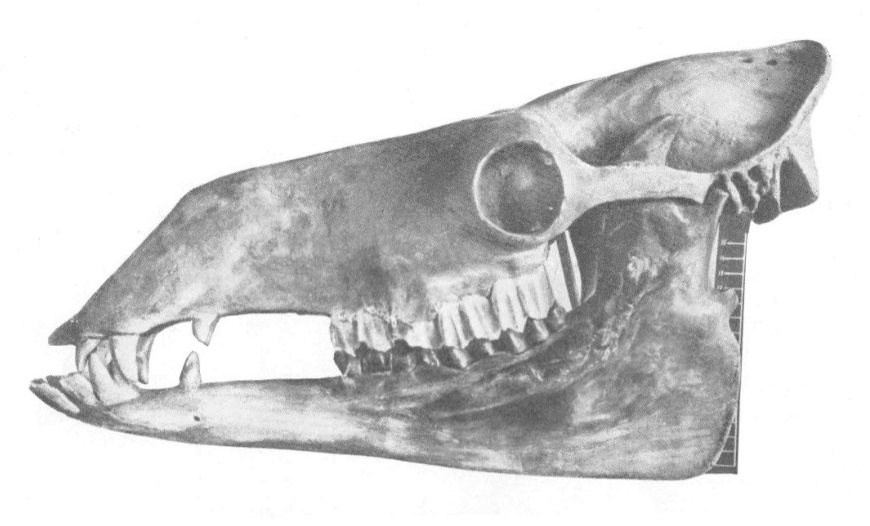
Череп с нижней челюстью паракамелюса (Paracamelus alexejevi), соединенный из частей от разных индивидов и в значительной части реконструированный, из позднего плиоцена Одесских катакомб в Одесской области, Украина.
По сравнению с Camelus, череп более узкий и длинный, особенно в лицевом отделе. Длина лицевого отдела составляет приблизительно 3/4 от основной длины черепа, а высота морды примерно в 5 раз ее меньше. Предчелюстные кости несколько вздуты. Носовые кости короткие, относительно уплощенные. Носовое отверстие узкое. Глазницы закрытые, направлены вбок и несколько вперед. Передний край глазницы находится над передним краем М3. Слезная вырезка узкая и длинная. Подглазничное отверстие находится примерно над серединой М4. Скуловая дуга незначительно вентрально искривлена. Теменные, сагиттальный и затылочный гребни резко выражены. Затылочный гребень высокий, тонкий, значительно нависающий над затылочными костями. Сагиттальный гребень единый, невысокий, соединяется с затылочным. Общее очертание затылочной поверхности приближается к трапеции. Затылочные мыщелки вытянуты и заострены каудально. Твердое небо вогнуто на всем протяжении, особенно на уровне предкоренных. Нижняя челюсть также сравнительно длинее, чем у рода Camelus, и относительно ниже: ее высота под серединой m3 составляет 15,5-17 % ее длины от переднего края до заднего края m3. В целом нижняя челюсть относительно длиннее, чем у Camelus. Ее тело иногда вздутое, в других случаях (вероятно, у самцов) лингвально-лабиально уплощенное.
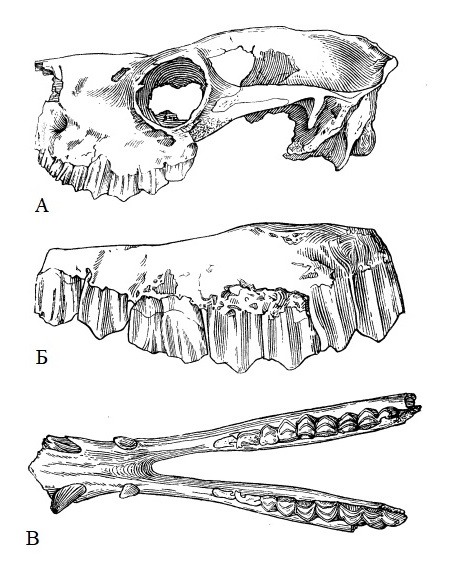
Прорисовки черепного материала паракамелюса (Paracamelus alexejevi) из позднего плиоцена Одесских катакомб в Одесской области, Украина: А — неполный череп; Б — левая верхняя челюсть с Р3-М3; В — нижняя челюсть, сверху.
Зубная формула: I1/i1, С1/с1, Р3/р3, M3/m3. Зубы мезогипсодонтные, сравнительно низкокоронковые. Аркады зубов располагаются почти параллельно сагиттальной плоскости черепа, образуя слабую дугу. Небольшой клыкообразный I3 расположен на предчелюстной кости, на небольшом расстоянии от клыка. Клыки развиты хорошо, овальные в сечении овальные. Редукция предкоренных выражена меньше, чем у рода Camelus. У самцов почти всегда, а у самок часто имеются клыкообразные P1/р1. Небольшой Р1 клыкообразный, отделен от клыка и от сомкнутых остальных щечных зубов большими диастемами. Р2 отсутствует. Р4 достаточно крупный и сильно моляризован. р3 хорошо развит; у молодых имеется pd2. р4 c открытой передней долинкой и с дополнительным вертикальным желобком в передне-внутреннем углу (этого желобка нет у близкого рода Gigantocamelus). Верхние коренные с хорошо развитыми парастилем, мезостилем и срединными ребрами на наружных поверхностях полулуний. m3 относительно короткий: его длина составляет 14-16 % длины челюсти до заднего края m3. На внешней стороне предкоренных и коренных зубов выражены вертикальные гребни, развитые сильнее, чем у рода Camelus. На m2 и m3 в большинстве случаев имеются передне-наружные складки.
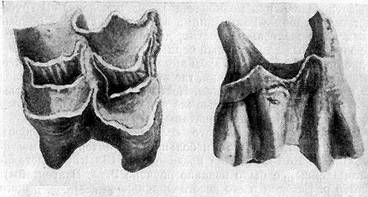
Коренной зуб паракамелюса (Paracamelus gigas) из раннего плейстоцена местонахождения Селим-Джевар на севере Казахстана.
Анатомически паракамелюсы сходны с представителями рода Camelus, но в большинстве случаев изящнее и мельче последних. Они обладали относительно стройными грацильными конечностями и шеей, что в совокупности с удлиненной лицевой частью черепа и относительно низкокоронковыми зубами показывает, что эти копытные сочетали питание в нижнем и верхнем растительных ярусах. По-видимому, паракамелюсы питались сравнительно мягким лиственным и веточным кормом. Для их метаподий характерны относительно меньшая ширина кости и более высокое расхождение в стороны суставных поверхностей дистального эпифиза по сравнению с таковыми у Camelus. Эти признаки могут свидетельствовать об ином способе локомоции данных животных по сравнению с современными верблюдами. Биомеханика позвоночника также отличалась от таковой у Camelus и в общих чертах ближе к роду Lama.
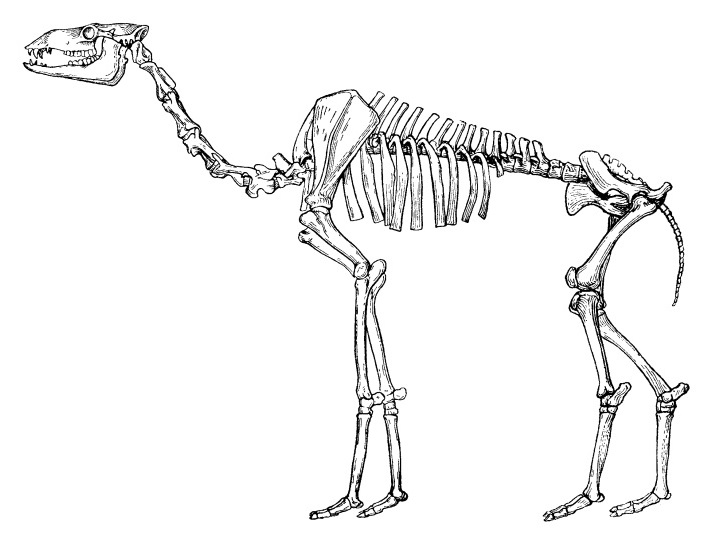
Прорисовка сборного скелета паракамелюса (Paracamelus alexejevi) из позднего плиоцена Одесских катакомб в Одесской области, Украина.
Состав
Род включает порядка 8 видов, большинство из которых описаны по фрагментам нижних челюстей.
Paracamelus aguirrei Morales, 1984 — вид, известный из позднего миоцена - раннего плиоцена (MN 13, мессинский - занклский ярусы) Приазовья, юга Украины, Крыма, севера Турции и др. Древнейший надежно установленный представитель рода.

Фрагмент нижней челюсти паракамелюса (Paracamelus cf. aguirrei) с p4-m3 из позднего миоцена Одессы в Одесской области, Украина: a — снаружи; b — изнутри.
Paracamelus longipes (Aubekerova, 1974) — вид, известный из плиоцена Казахстана. Синоним: Gigantocamelus longipes Aubekerova, 1974. Судя по строению черепа, зубов и конечностей, был приспособлен к существованию в условиях увлажненных ландшафтов.
Paracamelus praebactrianus (Orlov, 1927) Haveson, 1954 — вид, известный из позднего плиоцена - раннего плейстоцена Северного, Центрального и Восточного Казахстана, Таджикистана, Прииртышья, Западной Сибири и Северного Кавказа. Синоним: Camelus praebactrianus Orlov, 1927. Был описан Ю.А. Орловым по результатам сборов на р. Бетеке, Селим-Джевара, Кокчетавская область, Казахстан. Предложенное исследователем видовое название подчеркивает близость этого верблюда к двугорбому, однако без указания на предочность к нему. Легко сложенный, среднего размера верблюд, населявший открытые пространства.

Череп с нижней челюстью паракамелюса (Paracamelus praebactrianus) из Северного Казахстана.
Paracamelus gigas Schlosser, 1903 (тип) — типовой вид, известный из позднего плиоцена - среднего плейстоцена Северного Китая, Монголии, Северного и Восточного Казахстана, Таджикистана, Азербайджана, Приазовья (Ставропольский край, Ростовская область), юга Украины и Крыма. Описан в 1903 г. О. Шльоссером по фрагментарным остаткам, найденным на северо-востоке Китая. Самый крупный и рослый представитель рода: общая длина достигала 3,5 м, высота в холке — 2,4 м, вес оценивается в 0,8-1,4 т. Кости конечностей относительно массивные. Обитал в более аридных условиях, чем Paracamelus longiopes, т. е. во время поредения лесов и ксеротифизации растительного покрова.

Фрагмент нижней челюсти паракамелюса (Paracamelus gigas) из раннего плейстоцена местонахождения Винодельное в Ставропольском крае, Россия.
Paracamelus trofimovi Sharapov, 1986 — вид, известный из раннего плейстоцена Куруксая на юге Таджикистана. Размеры сравнительно небольшие.
Paracamelus khersonensis Pavlow, 1903 — вид, известный из позднего плиоцена юга Украины, Молдавии и Румынии. Стройный, средних размеров верблюд, достигавший величины небольшого дромадера (Camelus dromedarius). Иногда в рамках данного таксона рассматривают 2 хроноподвида — более древний Paracamelus khersonensis khersonensis и сменивший его Paracamelus chersonensis alexejevi.

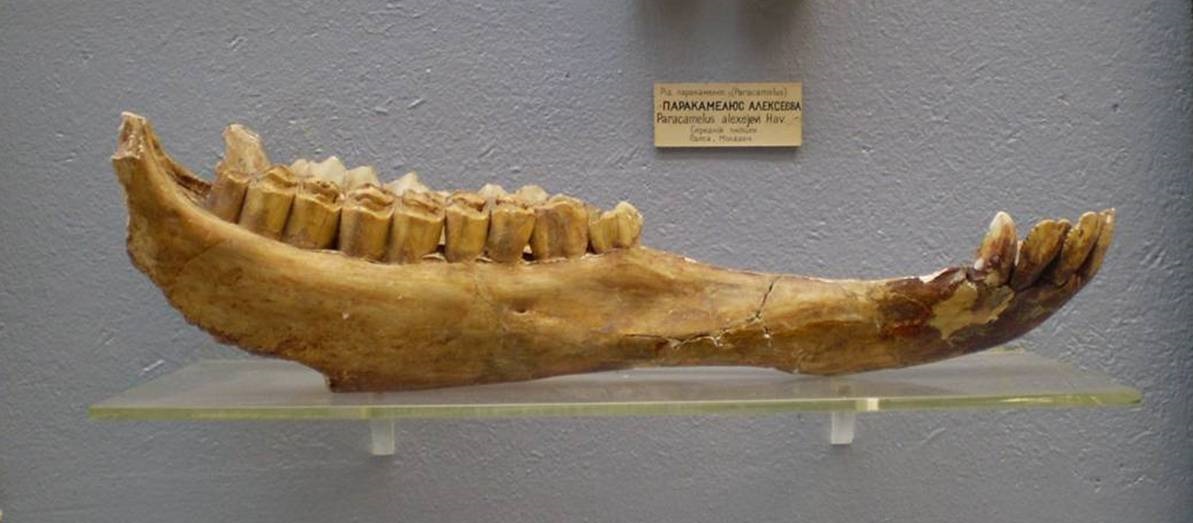
Неполный череп и нижняя челюсть паракамелюса (Paracamelus alexejevi) из позднего плиоцена Одесских катакомб в Одесской области, Украина.
Paracamelus alexejevi Havesson, 1950 — вид, известный из позднего плиоцена (MN 15 - MN 16, пьяченцский ярус) юга Украины и Молдавии. Является непосредственным потомком Paracamelus khersonensis. Найдено большое количество ископаемых остатков. Они преобладают в сборах из карстовых полостей известняков города Одессы (Одесских катакомб): здесь встречены части скелетов не менее 400 особей. Правда, ни один целый скелет не обнаружен, но более 50 тыс. костей, хотя и разрозненных, дают о животном отчетливое представление. Также найдены кости со следами заболеваний, патологических разращений в области суставов и т. д. Очевидно, этот верблюд был многочислен в Северном Причерноморье и при своей относительной беззащитности легко становился добычей крупных хищников.



Сборный скелет паракамелюса (Paracamelus alexejevi) из позднего плиоцена Одесских катакомб в Одесской области, Украина.
Ранее также указывалось на наличие вида Paracamelus minor Logvynenko, 2001, ошибочно описанного по фрагменту нижней челюсти Paracamelus alexejevi с молочными зубами из позднеплиоценовых отложений Одесских катакомб.
Paracamelus alutensis Stefanescu, 1895 — вид, известный из позднего плиоцена - раннего плейстоцена (MN 15 - MN 16, пьяченцский - гелазский ярусы) Приазовья, Причерноморья и сопредельных территорий. Очевидно, представляет собой измельчавшего потомка Paracamelus alexejevi. Животное достигало размеров современной ламы (Lama glama), приближаясь к наиболее мелким экземплярам одногорбого верблюда (Camelus dromedarius).
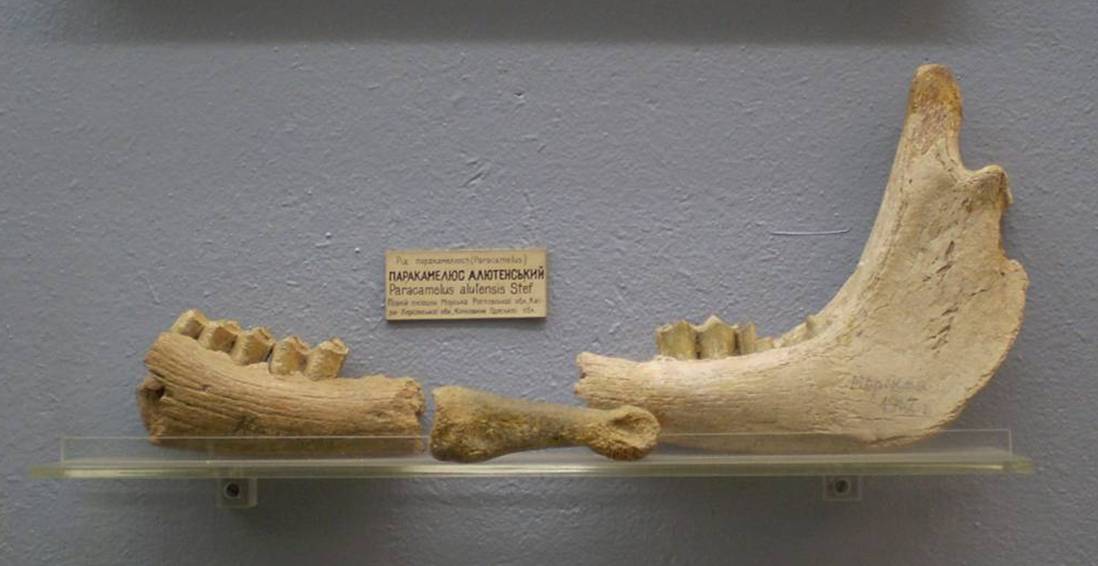
Части скелета паракамелюса (Paracamelus alutensis) из раннего плейстоцена местонахождения Морская в Ростовской области, Россия, а также местонахождений Каиры в Херсонской области и Котловина в Одесской области, Украина.
Иногда вид Paracamelus alutensis выделяют в отдельную подродовую группу неопаракамелюсов (Neoparacamelus Нavesson, 1954). В качестве основных признаков подрода Neoparacamelus принимаются относительно более мелкие размеры и отсутствие передне-наружных складок на m2 и m3. Однако имеющиеся данные показывают, что наличие складок — очень вариабельный признак, а мелкие размеры представителей данного вида не могут служить основанием для выделения отдельного подрода.

Позвоночный столб паракамелюса (Paracamelus alutensis) из раннего плейстоцена местонахождения Ливенцовка в Ростовской области, Россия.
Весьма примечателен фрагмент плюсневой кости Paracamelus alutensis со следами рубки и пиления-резания каменным орудием. Кость датируется финалом среднего виллафранка (2,1-1,97 млн. л. н.) и была обнаружена в Ливенцовском карьере (местонахождение Ливенцовка) на западной окраине Ростова-на-Дону вместе с другими фаунистическими остатками в хапровской аллювиальной толще, относящейся к русловой фации палео-Дона. Это одно из древнейших доказательств присутствия человека за пределами Африканского континента.

Дистальный фрагмент правой плюсневой кости паракамелюса (Paracamelus alutensis) из раннего плейстоцена местонахождения Ливенцовка в Ростовской области, Россия.
Другой мелкий представитель рода — Paracamelus kujalnensis Khomenko, 1915, — по всей вероятности, является младшим синонимом Paracamelus аlutensis. Признаки нижней челюсти из терминального раннего плейстоцена (куяльницкая формация) Причерноморья, на основании которых был выделен этот вид (меньшая относительная длина симфиза, несколько большая относительная высота горизонтальной ветви нижней челюсти и разница размеров), вероятно, являются результатом половой и индивидуальной изменчивости.
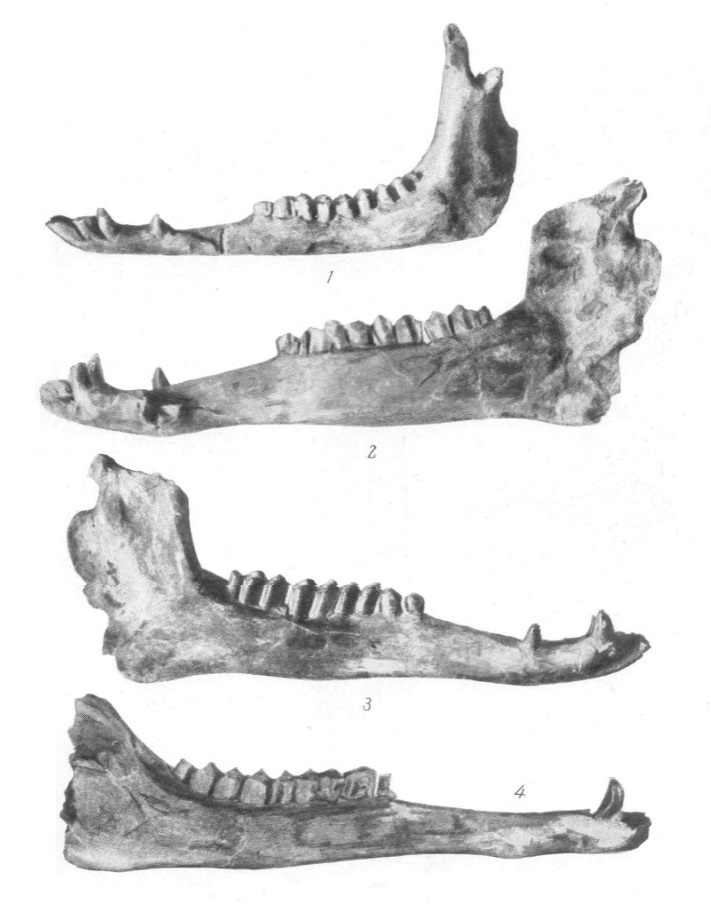
Нижние челюсти паракамелюсов: 1 — Paracamelus alutensis, снаружи; 2, 3 — Paracamelus alexejevi, изнутри и снаружи; 4 — Paracamelus kujalnensis, снаружи.
Остатки, определенные лишь до рода и обозначенные Paracamelus spp., известны из среднего миоцена США (штат Невада), позднего плиоцена Канады (территория Нунавут), раннего плиоцена Китая (провинция Шаньси), Казахстана и Болгарии, плиоцена Таджикистана и Украины (Одесская область), раннего плейстоцена Китая (провинция Хэбэй), раннего плейстоцена Турции и позднего миоцена Чада.
Paracamelus arenicola Reed et Longnecker, 1932 в настоящее время признается таксоном nomen nudum.

Частичный скелет паракамелюса (Paracamelus alutensis).
Эволюционная история
К Paracamelus наиболее близки такие североамериканские формы, как Gigantocamelus и Megatylopus. Самые древние известные остатки представителей рассматриваемого рода, обозначенные Paracamelus sp., происходят из среднего миоцена (формация Эмиеральда, кларендонский ярус) штата Невада, США. В Евразию паракамелюсы мигрировали через Берингийскую сушу в позднем миоцене, во времена повсеместного понижения уровня моря. Таким образом, эти верблюды стали первыми в Старом Свете представителями своего семейства. Удачная морфоанатомическая конструкция и распространение подходящих жизненных условий позволили им за достаточно короткое время широко расселиться в Восточном полушарии — в позднем миоцене паракамелюсы уже были многочисленны на гигантской территории от северной части Китая до северного Средиземноморья, откуда незадолго до мессинского кризиса солености в конце миоцена проникли на Аппенинский (Италия) и Пиренейский (Испания) полуострова, а не позднее начала плиоцена — и в Африку (Тунис, Кения, Чад).
Ранее при палеогеографических реконструкциях наличие в списке ископаемой фауны представителей рода Paracamelus рассматривался в качестве аргумента в пользу существования на данной территории пустынных или полупустынных ландшафтов, высоких температур и дефицита влаги. Однако внимательный анализ морфологии паракамелюсов и состава ископаемых фаун позволяет опровергнуть это мнение. По всей видимости, паракамелюсы являлись типичными обитателями сравнительно мезофитных степных и лесостепных ландшафтов, широко представленных в Евразии начиная с позднего миоцена. Косвенным подтверждением этому может быть тот факт, что типичными местообитаниями для многих североамериканских представителей Camelinae (Aepycamelus, Hemiauchenia, Alforjas, Paleolama, Camelops, Procamelus и Megatylopus) были открытые ландшафты саванного типа.
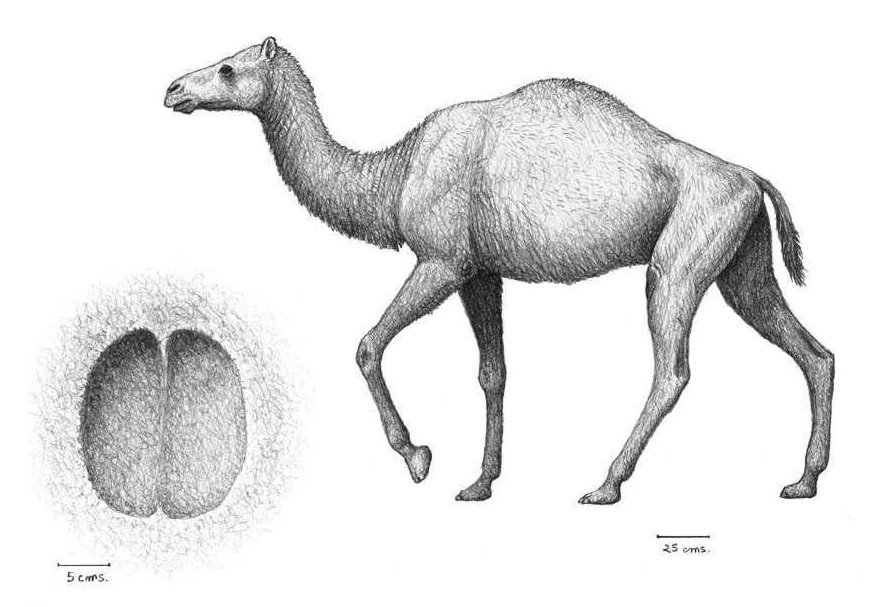
Реконструкция внешнего облика Paracamelus и его следовой отпечаток. Автор — М. Антон.
Позднемиоценовые и плиоценовые паракамелюсы являлись типичными представителями гиппарионовых фаун, которые в равной мере состояли из обитателей степей и лесостепей. На территории, простиравшейся от Центральной Азии до Африки, Paracamelus сосуществовали с мастодонтами, носорогами, гиппарионами, антилопами, жирафами и мелкими оленевыми. В позднем плиоцене и раннем плейстоцене по мере усиления аридизации открытых пространств изменился родовой состав фаун. На смену гиппарионовым ассоциациям пришли слоны, однопалые лошади, крупные оленевые и бычьи.
Для евразийских паракамелюсов предполагается существование 2 эволюционных линий. Первая последовательность включает Paracamelus aguirrei — Paracamelus сhersonensis — Paracamelus alexejevi — Paracamelus alutensis. Развитие этих верблюдов было связано, в первую очередь, с территорией Средиземноморья, включая Причерноморье и смежные территории. В данном регионе можно проследить постепенные эволюционные изменения паракамелюсов с позднего миоцена до конца позднего плиоцена. Остатки паракамелюсов здесь обычны в целом ряде местонахождений. В плиоцене большинство представителей этой линии вымерло на территории Африки и большей части Азии ввиду нарастания аридизации и остепнения. Более длительное время данная группа просуществовала на территории Причерноморья, где, возможно, дольше сохранялись отдельные стации и условия существования, характерные для предыдущих эпох.

Реконструкция предполагаемого образа жизни Paracamelus (по современным данным, неверная). Автор — К. Флеров.
История второй линии Paracamelus была тесно связана с Западной и Центральной Азией. Эти верблюды, в большей степени обладавшие адаптациями к существованию в открытых аридных ландшафтах, начали расселяться по Евразии вместе с распространением степей во второй половине плиоцена. Местами эти верблюды сосуществовали с представителями первой, "лесостепной", волны паракамелюсов.
По-видимому, именно один из представителей азиатской эволюционной линии еще до конца плиоцена стал предковым для рода Camelus. Можно предположить, что его появление было связано с увеличением аридных пространств на территории Евразии. Сами паракамелюсы исчезли до конца раннего плейстоцена, возможно, не выдержав повсеместного похолодания, либо были вытеснены из ранее типичных для них местообитаний оказавшимися более конкурентоспособными обитателями лесостепей и степей — крупными оленями и бычьими. Появление и массовое распространение этих животных происходило как раз в конце плиоцена и начале плейстоцена — во время вымирания Paracamelus и появления крупных форм Camelus.
Особый интерес представляют сильно фрагментированные остатки крупного верблюда, обозначенные Paracamelus sp. и происходящие из позднего плиоцена (около 3,4 млн. лет назад) канадского острова Элсмир, расположенного за полярным кругом к западу от северного побережья Гренландии. В то время средняя температура на Элсмире составляла -1,4 °C, что примерно на 18 °C выше нынешних показателей. Остров был покрыт редколесьем, перемежавшимся торфяниками. Он лежит на 1200 км севернее тех мест, где до этого находили остатки древних верблюдов. Таким образом, арктические территории также могли сыграть свою роль в становлении рода Camelus.
Выработанные современными верблюдами приспособления к жизни в засушливых условиях (горбы, особенности пищеварения и обмена веществ, поглощение большого количества воды и в связи с этим потребность в кормах, содержащих много соли, иноходь и пр.), вероятно, появились в эволюции верблюдовых относительно поздно и не были характерны для них изначально. Неприхотливость видов Camelus явилась причиной распространения этих необычных животных в условиях, непригодных для других травоядных.
Ссылки
Паракамелюсы (Википедия) перейти
Paracamelus (Paleobiology Database) перейти
Литература
Кожамкулова Б. С. Позднекайнозойские копытные Казахстана. — Алма-Ата: Наука, 1981
Кожамкулова Б. С., Костенко Н. Н. Вымершие животные Казахстана (Палеогеография позднего кайнозоя). — Алма-Ата: Наука, 1984
Орлов Ю. А. В мире древних животных. 3-е изд. — М.: Наука, 1989
Основы палеонтологии. В 15 т. Т. 13: Млекопитающие / Гл. ред. Орлов Ю. А. — М.: Государственное научно-техническое издательство литературы по геологии и охране недр, 1962
Титов В.В. Крупные млекопитающие позднего плиоцена Северо-Восточного Приазовья. — Ростов-на-Дону: Изд-во ЮНЦ РАН, 2008
Титов В.В. Особенности морфологии и экологии верблюдов рода Paracamelus // Современная палеонтология: классические и новейшие методы. — М.: ПИН РАН, 2005
Augusti J., Anton M. Mammoths, sabertooths, and hominids: 65 million years of mammalian evolution in Europe. — New York: Columbia University Press, 2002