Пеликозавры — Pelycosauria
Клада Амниоты (Amniota)
Клада Синапсидообразные (Synapsidomorpha)
Класс †Синапсиды (Synapsida)
Подкласс †Пеликозавры (Pelycosauria)
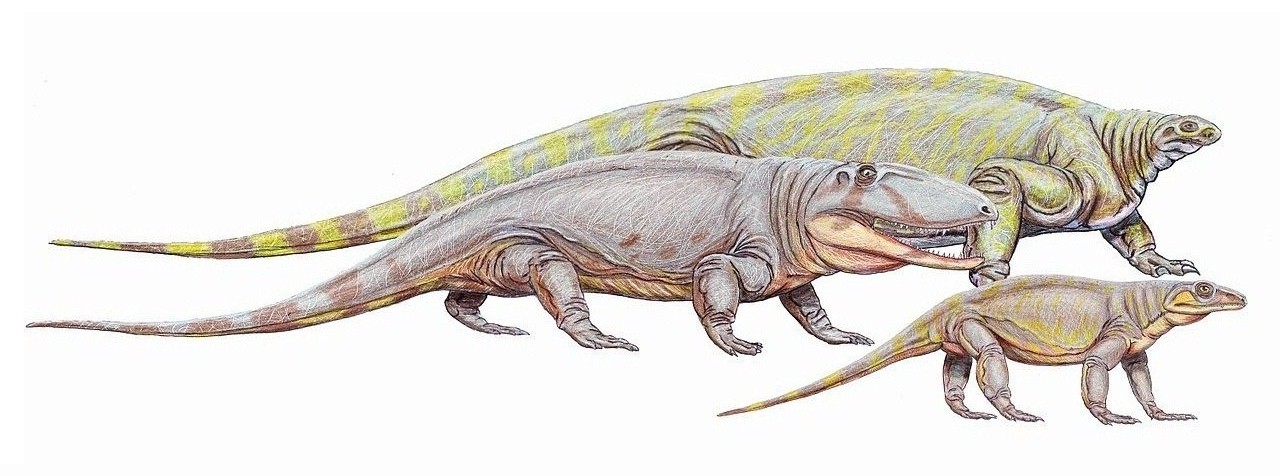
Пеликозавры (Pelycosauria Cope, 1878) — в классическом понимании группа, объединяющая низших синапсид; условно принимается здесь в качестве подкласса. Являются парафилетической группой, поэтому сторонники кладистической классификации не считают ее таксоном, а название группы берут в кавычки: "пеликозавры". В целом сохраняют типично рептильный внешний облик и физиологию. Одна из ветвей является предковой для терапсид. Выделено от 6 до 10 семейств и примерно 50 родов пеликозавров.
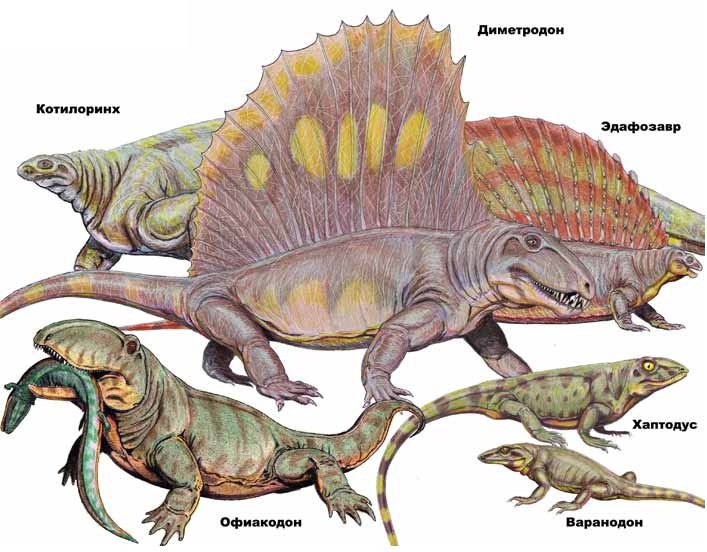
Разнообразие пеликозавров. Автор — Д. Богданов.
Название группы можно перевести как "ящеры в шлемах", что вызвано строением черепа данных животных.
Возраст и распространение
Пеликозавры известны со среднего карбона до средней перми. Их ископаемые остатки найдены преимущественно в Северном полушарии (Европа, Азия и Северная Америка), но обнаружены также на юге Африки. Неописанные остатки известны из Южной Америки. Особенно обильны остатки пеликозавров в США (штаты Техас, Оклахома, Нью-Мексико) и в Западной Европе (Чехия, Франция, Германия). На территории бывшего СССР их обломочные остатки встречены в раннепермских отложениях Приуралья и Казахстана, а скелеты хорошей сохранности — в среднепермских отложениях Архангельской области России.
Строение и функции
Пеликозавры достигали приблизительно от 0,5 до 6 м в длину. Внешне большинство из них походило на крупных ящериц. В целом эти синапсиды находились на уровне организации, характерном для ранних амниот, и морфологически во многом близки к капториноморфам.
Череп обычно высокий и узкий, но бывает также низким и широким. Морда от сильно удлиненной до укороченной. Ноздри широко расставлены. У большинства форм, кроме высших сфенакодонтид, слезные кости удлиненные и зачастую достигают края ноздрей. Септомаксилла небольшая или хорошо развитая. У казеид, в частности, у Ennatosaurus, необычно массивная септомаксилла закрывает заднюю часть ноздри, по верхнему краю которой широко сочленяется с носовой костью. У эотиридида Oedaleops септомаксилла полностью закрывает нижний край очень длинной ноздри. У эдафозаврид она погружена глубоко в ноздрю и соприкасается со слезной и носовой костями, будучи отделена от этих костей отверстиями. У многих сфенакодонтид, например, у Dimetrodon, септомаксилла развита по всему заднему краю ноздри и достигает наверху носовой кости, образуя вдающийся в ноздрю передний "дерминтермедиальный" отросток. От максиллы — а у примитивнейшего сфенакодонтида Haptodus также от слезной кости — септомаксилла отделена отверстиями.
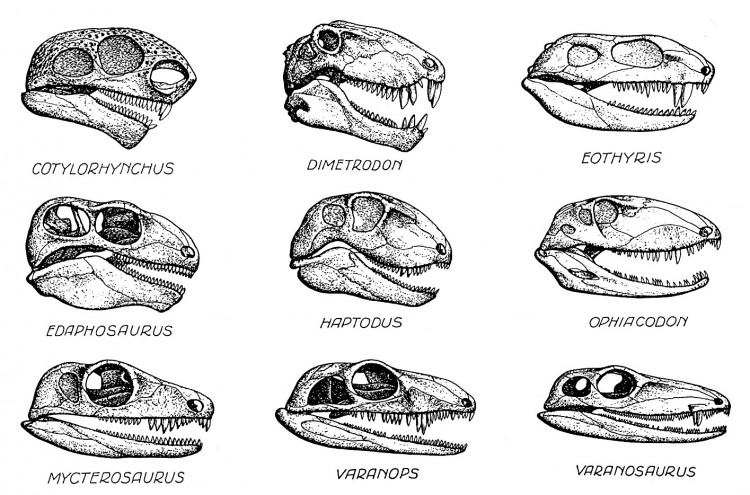
Прорисовки черепов с нижними челюстями различных пеликозавров; взаимный масштаб не соблюден.
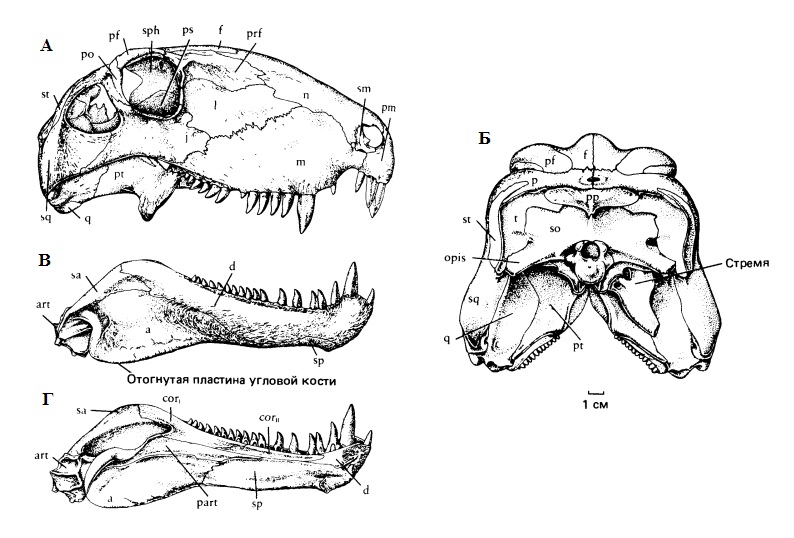
Типичные пеликозавры отличаются относительно короткой заглазничной областью. Височное окно обычно относительно небольшое и снизу ограничено скуловой и чешуйчатой костями, а сверху — чешуйчатой и заглазничной. Это отличает пеликозавров от терапсид. Но у варанопид височное окно увеличено в размерах, а квадратно-скуловая кость, как и у терапсид, входит в край височного окна. Как и у ранних диапсид, наличие височного окна, по-видимому, не сильно сказывалось на расположении челюстных мышц пеликозавров, хотя у терапсид оно имеет большое значение. Возможно, появление этого окна в щечной области улучшило распределение механических напряжений в задней части черепа. Обычно сохраняется маленькая надвисочная кость. Теменные кости широкие, не дают нисходящих отростков и отделены от переднеушной кости неокостеневающим пространством. Пинеальное отверстие обычно большое. Заднетеменная и таблитчатые кости обычно выходят на дорсальную поверхность черепа, но развиты главным образом на его затылочной поверхности. Затылок широкий и высокий, обычно сильно скошенный назад. Затылочный мыщелок одинарный и образован главным образом основной затылочной костью.
Прорисовки черепа с нижней челюстью продвинутого пеликозавра Dimetrodon: череп сбоку (А) и сзади (Г), нижняя челюсть снаружи (Б) и изнутри (В).
Вторичного неба нет. Сошник парный. Базиптеригоидные отростки хорошо развиты, базиптеригоидное сочленение подвижное, межптеригоидные ямы длинные и узкие. Боковые отростки крыловидных костей обычно хорошо развиты, и крыловидные кости соприкасаются со скуловыми. Квадратная ветвь крыловидной кости очень высокая и ограничивает спереди полость среднего уха. Верхняя крыловидная кость нерасширенная. Квадратная кость неподвижная. У офиакодонтид сохраняется самостоятельная верхнезатылочная кость, не соединенная швом с крышей черепа или прочно связанная либо с таблитчатой костью, либо со слуховой капсулой. Последняя мала и не соприкасается со щечной областью. У других пеликозавров кости затылка объединены в пластиновидную структуру, включающую верхнезатылочную кость, широкие парокципитальные отростки и краевые дермальные кости черепа. Слуховая косточка (стремя) массивная, с дорсальным отростком, идущим к околоушному отростку. Контактируя со щечной областью черепа, она, по-видимому, выполняет функцию его укрепления у ранних пеликозавров, но и в дальнейшем остается крупным элементом у всей этой группы, хотя после упрочнения затылка уже не играет структурной роли. Ушной вырезки нет. Челюстной сустав, образованный квадратной и сочленовной костями, отодвинут каудально, занимая положение позади затылочного мыщелка. Задние кости нижней челюсти не редуцированы, порой имеются 2 венечные кости. Угловая кость иногда с вырезкой.
Челюстные зубы сидят в альвеолах, конической формы, загнутые назад. Верхнечелюстные клыковидные зубы обычно более или менее дифференцированы, нижнечелюстные отсутствуют. У примитивных форм на птеригоидах хорошо развиты краевые зоны с небными зубами, которые также могут покрывать парасфеноид.
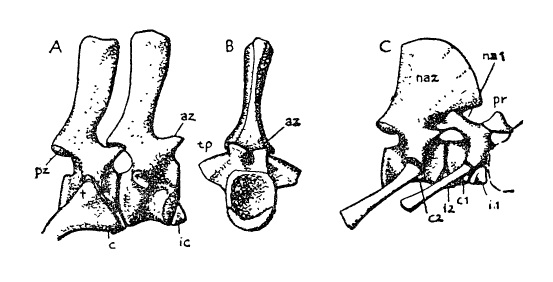
Прорисовки позвонков примитивного пеликозавра Ophiacodon. А, В — задние шейные позвонки, вид сбоку и спереди: аz — передняя зигапофиза; c — ребрная головка; ic — интерцентр; pz — задняя зигапофиза; t — реберный бугорок; tp — processus traversus. C — атлант и эпистрофей, вид сбоку: c1, c2 — тела атланта и эпистрофея; i1, i2 — интерцентры атланта и эпистрофея; na1, na2 — невральные дуги; pr — проатлант: невральная дуга, тело которой, по-видимому, вошло в состав основной затылочной кости; мыщелок черепа обозначен прерывистой линией.
Тела позвонков глубоко амфицельные, с отверстием для хорды. Невральные дуги уже, чем у капториноморф. Хорошо развитые интерцентры продолжают существовать на всем протяжении позвоночного столба. Обычно имеется 27 предкрестцовых, 2-4 крестцовых и около 60 хвостовых позвонков. Проатлант имеется. Остистые отростки позвонков изначально низкие и треугольные, но эдафозавров и некоторых сфенакодонтов они резко вытягиваются на спине, постепенно уменьшаясь к шее и хвосту. Поперечные отростки позвонков также удлиняются. Эти изменения, возможно, связаны с ранней стадией реорганизации туловищной мускулатуры. У млекопитающих развитие осевой мускулатуры над и под поперечными отростками помогает ограничивать боковые волнообразные движения туловища и поддерживать позвоночный ствол в виде дуги над поясми конечностей. Боковая ундуляция позвоночника остается важным элементом локомоции у большинства синапсид, но удлинение поперечных отростков, возможно, означает его более активную опору на пояса конечностей, чем у других ранних амниот. Ребра обычно двухголовчатые.
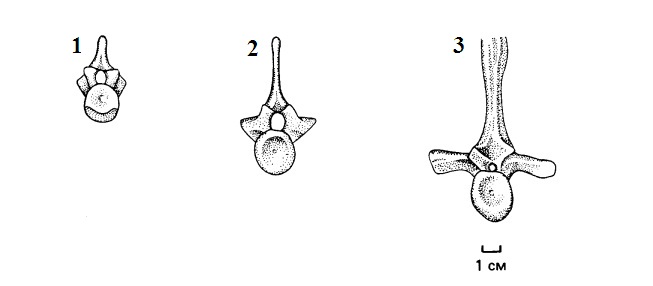
Прорисовки туловищных позвонков пеликозавров, спереди: 1 — Protoclepsydrops; 2 — Archaeothyris; 3 — Dimetrodon. Видно увеличение длины поперечных отростков со среднего карбона до ранней перми.
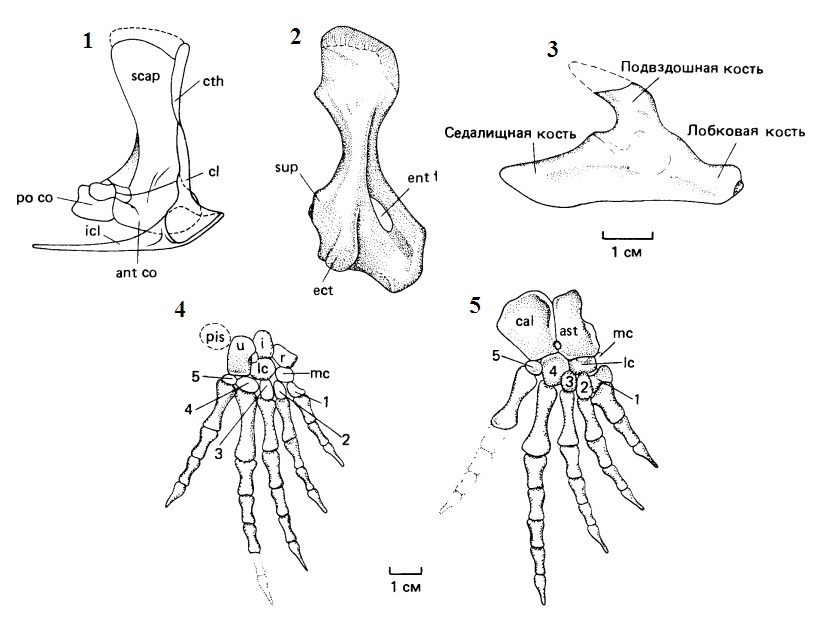
В плечевом поясе сохраняется клейтрум, который у терапсид обычно отсутствует. Лопатка широкая, без акромиального отростка. Суставная ямка плечевого сустава винтообразно изогнутая. Над суставной ямкой имеется утолщение, обычно прободенное отверстием. Задний коракоид невелик и варьирует по своему строению. Ключицы вентрально несколько расширены; межключица с задним отростком. Подвздошная кость направлена вверх и назад и обычно не разрастается вперед от вертлужной впадины. Вентральный отдел тазового пояса пластинчатый, тироидного отверстия нет. Суставная ямка бедренноого сустава также имеет винтообразную форму. У ранних пеликозавров имеются лишь 2 крестцовых позвонка и узкая подвздошная кость. У некоторых групп продвинутых пеликозавров тазовое сочленение усилено 1 или 2 дополнительными крестцовыми позвонками, а пластина подвздошной кости широкая для поддержания более тяжелого тела.
Прорисовки элементов конечностей и их поясов пеликозавров: 1 — плечевой пояс Dimetrodon, демонстрирующий тройное разделение скапулокоракоида. 2 — плечевая кость Archaeothyris. Наличие выступающего супинаторного отростка и эктэпикондилярного гребня типично для примитивных пеликозавров. Как и у всех ранних рептилий, имеется крупное энтэпикондилярное отверстие. У некоторых пеликозавров супинаторный отросток распространяется дистально, окружая энтэпикондилярное отверстие. 3 — таз Archaeothyris. 4, 5 — кисть и стопа Haptodus. Сокращения: ant со — передний коракоид; ast — таранная кость; cal — пяточная кость; cl — ключица; cth — клейтрум; ect — эктэпикондилярный гребень; ent f — энтэпикондилярное отверстие; i — промежуточная кость; icl — межключица; lc — латеральная централия; mc — медиальная централия; pis — гороховидная кость; ро со — задний коракоид; r — радиалия; scap — лопатка; sup — супинаторный отросток; u — ульнария; 1-5 — дистальные карпалии и тарсалии.
Конечности по сравнению с капториноморфами длиннее и тоньше, задние обычно длиннее передних. Кости конечностей массивные, с крупными буграми для крепления мышц. Подобно амфибиям, плечо и бедро располагаются в горизонтальной плоскости (латеральное положение конечностей). Плечевая кость с резко расширенными концами и винтообразной проксимальной сочленовной головкой. Особенно сильно расширен дистальный отдел плеча, где над радиальным надмыщелком развит массивный энтэпикондилярный, или супинаторный, отросток. Головки плечевой кости расположены под значительным углом друг к другу. Энтэпикондилярное отверстие хорошо выражено, но эктэпикондилярное обычно отсутствует. Проксимальная головка бедренной кости не смещена внутрь от длинной оси последней. Тело бедренной кости несет обычно хорошо выраженный IV-й трохантер. При перемещении движения передней конечности сопровождались поворотом предплечья относительно плеча, а движения задней конечности были упрощены, хотя их тоже сопровождал поворот голени относительно бедра. В кисти и стопе сохраняются по 2 центральных косточки. Фаланговая формула, за редкими исключениями (Caseidae), обычная для рептилий — 2 - 3 - 4 - 5 - 3 или 4. Как правило, имеются гастралии.
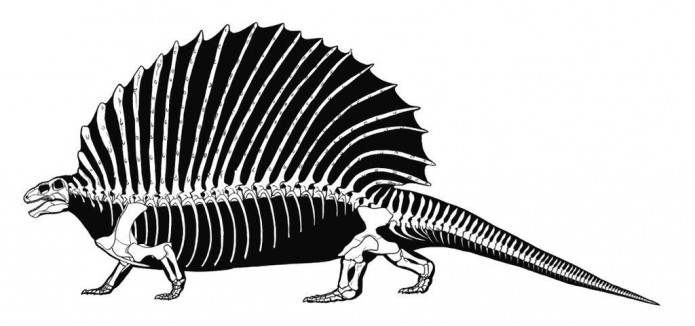
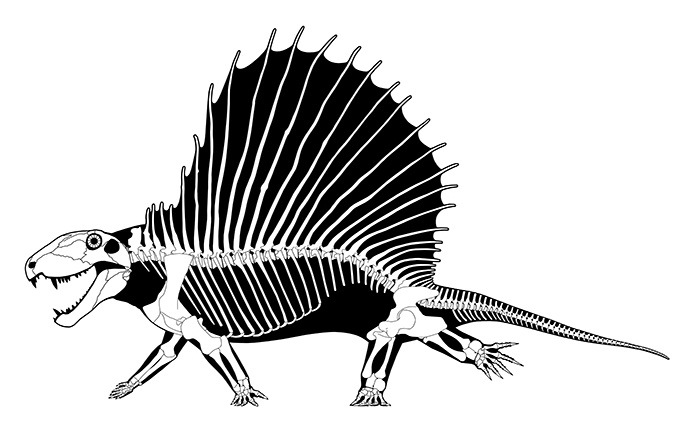
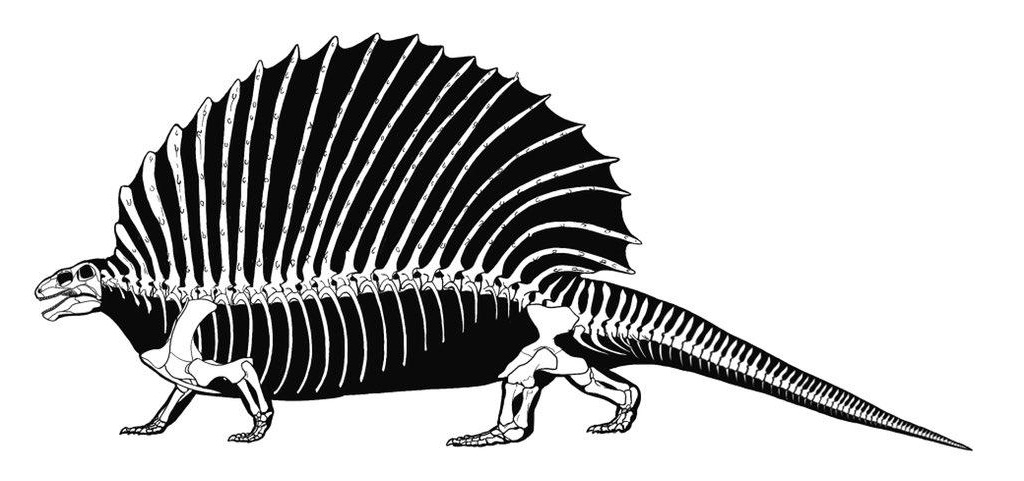
Прорисовки реконструированных скелетов прогрессивных пеликозавров эдафозавра (Edaphosaurus cruciger) и диметродона (Dimetrodon grandis), вписанных в телесные очертания.
Судя по сохранившимся следовым отпечаткам, пеликозавры ходили приподняв туловище над землей. Когда отдыхали — ложились на землю всем телом (сохранились соответствующие отпечатки). Однако никто из них не ползал — борозды от волочащегося брюха никогда не обнаруживаются. Судя по всему, передвижение было довольно медленным, особенно у крупных животных.


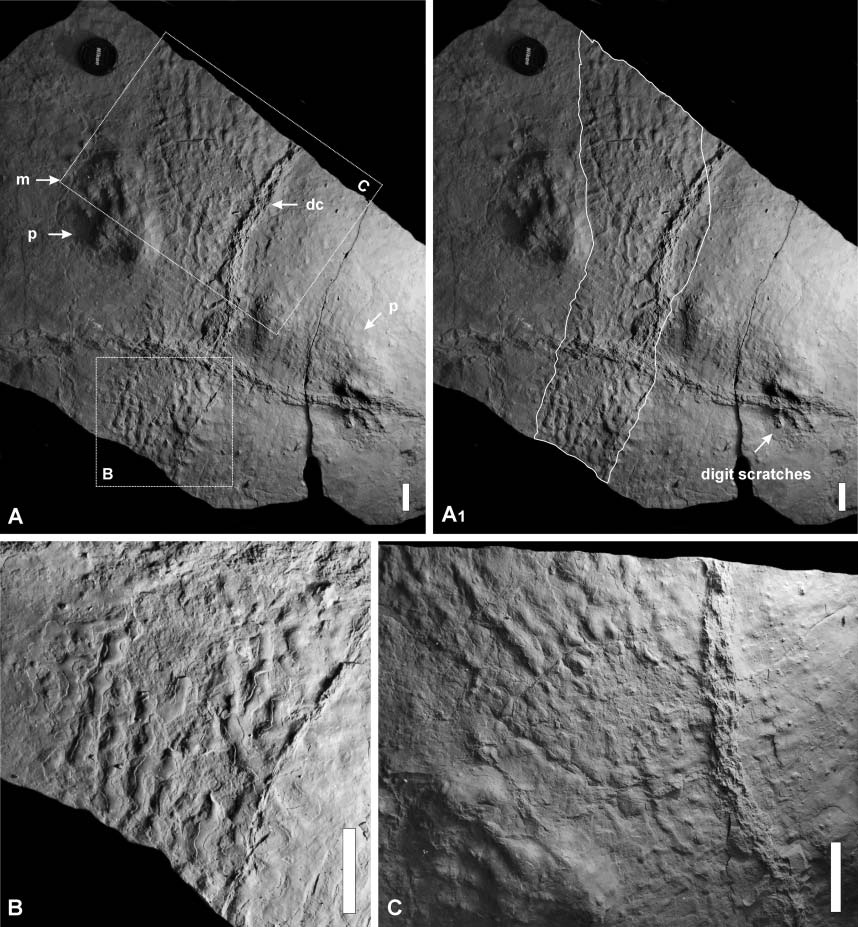
Следовые отпечатки ихнотаксона Dimetropus leisnerianus из ранней перми (низы ассельского яруса, приуральский отдел) Босковицкого бассейна, Чехия, и гипотетическая реконструкция внешнего облика оставившего их сфенакоморфа.
Предполагают, что кожа пеликозавров была гладкой, с немногочисленными кожными железами. Иногда на верхней стороне тела присутствуют мелкие кожные окостенения. В раннепермских отложениях Польши найден отпечаток нижней стороны тела отдыхавшего в грязи пеликозавра, при этом брюхо и основания лап покрыты чешуями разного размера. Однако этот отпечаток может принадлежать офиакодонту, находящемуся в стороне от синапсидной линии. Также не исключено, что "чешуи" — это просто отпечатки, оставшиеся, когда тело животного отлепилось от вязкой грязи.

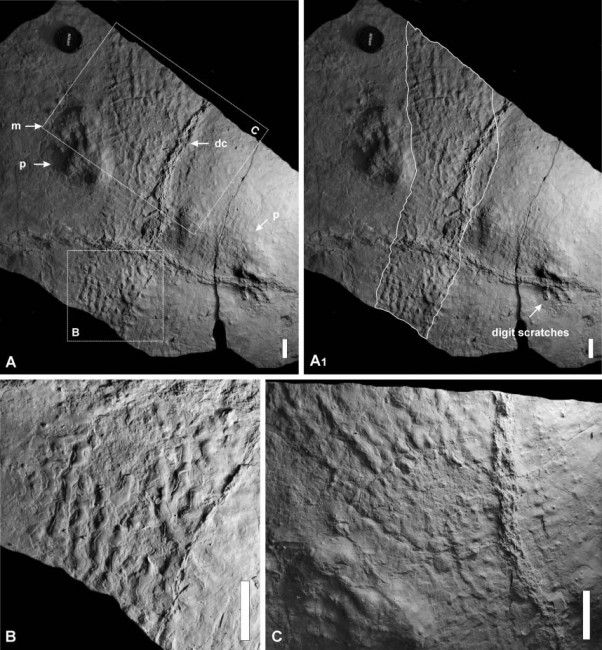
Отпечатки брюха и лап ихнотаксона Dimetropus leisnerianus из ранней перми Польши. Длина масштабной линейки — 5 см.
Не выявлено никаких достаточно убедительных признаков, предполагающих какую-либо тенденцию к повышению интенсивности метаболизма или формированию иных прогрессивных физиологических характеристик, типичных для млекопитающих. По своей физиологии пеликозавры были, по-видимому, сходны с современными рептилиями типа ящериц, черепах и крокодилов. По утрам они грелись на солнце, а в жару прятались в тень. Впрочем, пеликозавры могли являться и преимущественно ночными животными. С другой стороны, гистологические исследования костей примитивных пеликозавров Ophiacodon указывают на их быстрый, хотя и с перерывами, рост. Таким образом, росли они скорее как млекопитающие, чем как рептилии.
Другой признак возможного наличия температурного контроля у ряда родов — высокие остистые отростки туловищных позвонков, порой вытянутые в длину более чем на 1 м. Хорошая их сочлененность в ископаемом состоянии, даже когда отдельные отростки сломаны, говорит о том, что они были заключены в единую пластину мягкой ткани наподобие паруса. Замечено, что длина остистых отростков у Dimetrodon возрастает аллометрически по сравнению с другими линейными размерами, так что площадь этого "паруса" увеличивается пропорционально объему тела. В бороздах у основания отростков, по-видимому, проходили кровеносные сосуды. Васкуляризация "паруса", возможно, позволяла использовать его для быстрого поглощения или отдачи тепла телом. Животное поворачивалось боком к солнцу и кровь прогревалась, проходя через сосуды в "парусе", а в глубокой тени либо на ветру она охлаждалась.
Хотя наличие "паруса" может указывать на отбор, благоприятствующий терморегуляции, оно равным образом свидетельствует об эктотермной, а не эндотермной физиологии этих животных. По другим предположениям, "парус" использовался для распознавания сородичей, демонстраций в брачных ритуалах, служил для стабилизации позвоночника, маскировки среди вертикальных стеблей растений или в качестве настоящего паруса при плавании. У различных живущих по соседству видов "парус" мог быть окрашен в разные цвета или иметь специфический узор.
Косвенным свидетельством в пользу пойкилотермности пеликозавров также может служить качественный состав содержащих их фаунистических комплексов. Количеством хищников и растительноядных в них примерно одинаково, тогда как в гомойотермных сообществах хищников всегда меньше, чем их жертв.
Пеликозавры из ранней перми штата Техас, США. Слева направо изображены Casea, Edaphosaurus и Dimetrodon; в воде амфибии Diplocaulus. Автор — Ч. Найт, 1931 г.
Образ жизни
Пеликозавры вели наземный образ жизни; некоторые, возможно, были роющими животными. По характеру питания среди них выделяются плотоядные, насекомоядные и растительноядные формы.
В конце карбона и начале перми пеликозавры доминировали в экосистемах суши. Так, в позднем карбоне они составляли примерно половину всех известных родов амниот, а к началу пермского периода их доля возросла до 70 %.
Представители офиакоморф: спереди назад показаны Varanops, Ophiacodon и Cotylorhynchus. Автор — Д. Богданов.
Систематика
К сожалению, еще плохо известны ископаемые остатки из карбона, когда происходила первоначальная дифференциация пеликозавров, поэтому конкретные взаимосвязи различных их групп продолжают оставаться спорными.
А. Ромер и Л. Прайс (1940 г.) предложили исчерпывающий на тот момент анатомический и таксономический обзор Pelycosauria. Они различали 3 их подотряда: Ophiacodontia (семейства Ophiacodontidae и Eothyrididae), Sphenacodontia (семейства Varanopseidae и Sphenacodontidae) и Edaphosauria (семейства Edaphosauridae, Lupeosauridae, Nitosauridae и Caseidae). К офиакодонтам были отнесены примитивные предковые формы, известные начиная с конца среднего карбона, и относительно крупные, преимущественно рыбоядные, формы из нижней перми. Эдафозавры — это примущественно растительноядные пеликозавры, появившиеся в позднем карбоне, но особенно многочисленные в ранней перми. Плотоядные сфенакодонты преобладали в ранней перми; по наличию дополнительного наружного отростка (lamina reflecta) на угловой кости нижней челюсти они уже сходны с терапсидами. Со временем выявилась целесообразность выделения в особую группу, по-видимому, растительноядного семейства Caseidae, которое А. Ромер и Л. Прайс включили в эдафозавров, а также хищного семейства Varanopseidae, отнесенного ими к сфенакодонтам.
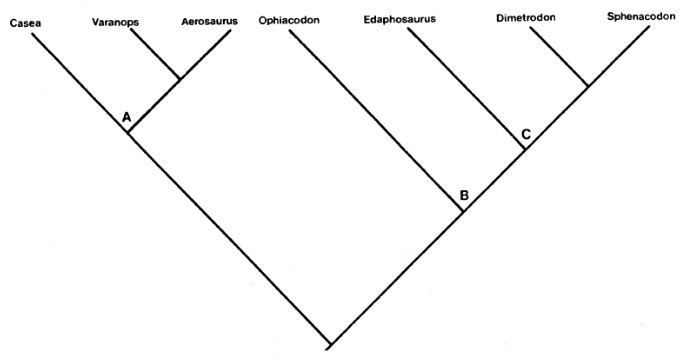
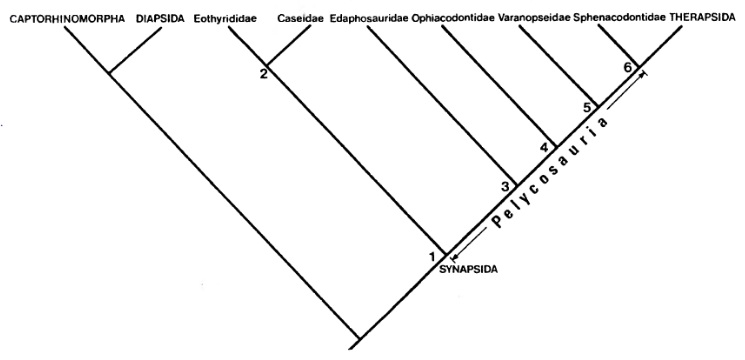
Д. Бринкман и Д. Эберт (1983 г.) поставили под сомнение валидность подотрядов, признав, однако, большинство семейств, выделенных А. Ромером и Л. Прайсом. Воззрения Д. Бринкмана и Д. Эберта выражает следующая кладограмма:
Р. Рейс (1987 г.) включил в отряд Pelycosauria 2 подотряда: Casesauria (семейства Eothyrididae и Caseidae) и Eupelycosauria (семейства Varanopsidae, Ophiacodontidae, Edaphosauridae и Sphenacodontidae). Кладограмма пеликозавров по Р. Рейсу выглядит так:
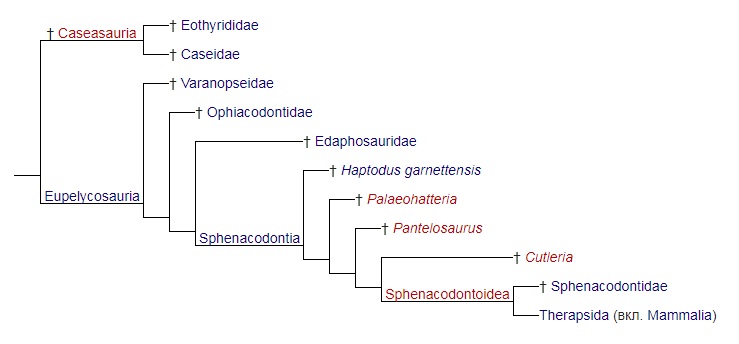
М. Лорин и Р. Рейс (1997 г.) пересмотрели таксон Eupelycosauria в духе кладистики для обозначения группы синапсид, которая включает большинство пеликозавров, а также всех терапсид и млекопитающих. Эту точку зрения иллюстрирует следующая кладограмма:
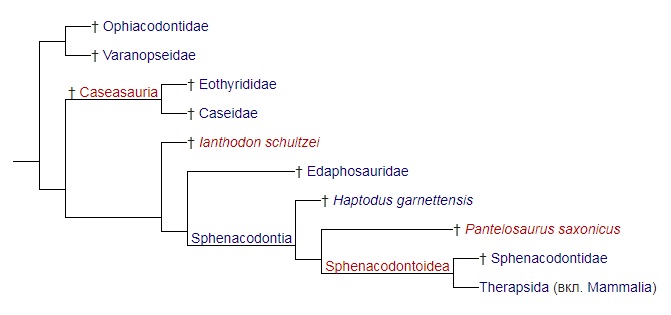
Однако по результатам структурного анализа 45 таксонов пеликозавров, выполненного Р. Бенсоном (2012 г.), филогенетические связи семейств пеликозавров представляются такой кладограммой:
Таким образом, по данным Р. Бенсона семейства Ophiacodontidae и Varanopseidae составляют кладу, которая ответвилась от филогенетического древа пеликозавров ранее казеазавров. Данные результаты ставят под сомнение обоснованность выделения подотряда эвпеликозавров в его ныне принятом объеме.
М.Ф. Ивахненко (2008 г.) вообще отрицает принадлежность семейств Caseidae, Eothyrididae, Ophiacodontidae и Varanopidae к синапсидной ветви амниот и считает необоснованным выделение как класса Synapsida, так и отряда Pelycosauria. Вместо этого он предлагает рассматривать в составе рептилий 2 новых подкласса, включая представителей указанных 4 семейств в один из них — подкласс Ophiacomorpha, а всех остальных синапсид (включая семейства Edaphosauridae и Sphenacodontidae) — в подкласс Theromorpha. Подкласс Ophiacomorpha включает в себя отряд Ophiacodontia (надсемейство Varanopoidea с семействами Varanopidae и Watongiidae, а также надсемейство Ophiacodontoidea с семейством Ophiacodontidae) и отряд Casesauria (семейства Eothyrididae и Caseidae).
Офиакоморфы характеризуются особенностями строения височного окна (оно ограничено квадратноскуловой, скуловой и чешуйчатой костями) и аддукторной полости (ее медиальная стенка сформирована пластиной палатоквадратума). Это отличает данную группу от истинных пеликозавров — сфенакодонтов и эдафозавров. Кроме того, офиакоморфы не обладают периангулярной полостью (и, соответственно, ангулярным лепестком) на угловой кости нижней челюсти. Таким образом, наличие синапсидного окна не является достаточным основанием для включения всех "синапсид" в единую группу. По мнению Ивахненко, Ophiacomorpha состоят в более близком родстве с Archosauromorpha, чем с Theromorpha, приобретя некоторые синапсидные признаки независимо от последних.
Как особая группа офиакоморфы признаются лишь отдельными исследователями и вопрос этот требует дальнейшего изучения.
Ссылки
Pelycosauria (Paleobiology Database)
перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Иванов А. О., Черепанов Г. О. Ископаемые высшие позвоночные. — СПб.: Издательство Санкт-Петербургского университета, 2001
Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1 / Отв. ред. Ивахненко М. Ф. и Курочкин Е. Н. — М.: ГЕОС, 2008
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Павлова М. В. Палеозоология. Ч. 2. Позвоночные. — М.-Л.: Государственное издательство, 1929
Ромер А. Ш. Палеонтология позвоночных. — М. — Л.: Государственное научно-техническое издательство нефтяной и горно-топливной литературы, 1939
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГЕОС, 2009