Плезиориктеропусы — Plesiorycteropus
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Афротерии (Afrotheria)
Клада Афроинсектифилии (Afroinsectiphilia)
Клада Афроинсективоры (Afroinsectivora)
Отряд Афросорициды (Afrosoricida)
Подотряд Тенрекообразные (Tenrecomorpha)
Надсемейство Тенрекоиды (Tenrecoidea)
Семейство Тенрековые (Tenrecidae)
Подсемейство †Плезиориктероподины (Plesiorycteropodinae)
Род †Плезиориктеропусы (Plesiorycteropus)
Вид †Plesiorycteropus madagascariensis
Вид †Plesiorycteropus germainepetterae
Плезиориктеропусы (Plesiorycteropus Filhol, 1895) — род монотипического подсемейства плезиориктеропин (Plesiorycteropodinae Patterson, 1975), происходящий из ? позднего плейстоцена - позднего голоцена Мадагаскара. Установлены 2 вида.

Реконструкция внешнего облика плезиориктеропуса (Plesiorycteropus madagascariensis). Автор — Alphynix.
Синонимы: Hypogeomys Grandidier, 1869 (частью); Myoryctes Forsyth Major, 1908, non Ebert, 1863; Majoria Thomas, 1915.
История изучения
Род и типовой вид Plesiorycteropus madagascariensis установил в 1895 г. французский натуралист Анри Фильоль по частичному черепу, обнаруженному в пещере Бело-сур-мер на юго-западном побережье Мадагаскара. Описание было расплывчатым даже по стандартам XIX в. и не содержало иллюстраций. Основываясь на общем морфологическом сходстве, ученый отнес открытое им животное к отряду трубкозубых (Tubulidentata) и семейству трубкозубовых (Orycteropodidae). Это отразилось в родовом наименовании: πλησίος (plesio) в переводе с древнегреческого языка означает "близкий", а Orycteropus — латинское название современного трубкозуба; видовой эпитет указывант на Мадагаскар как на место находки. Зачастую зверя называли просто мадагаскарским трубкозубом.
Первое время найденные остатки плезиориктеропусов несколько раз ошибочно приписывались грызунам и лемурам. Так, в 1908 г. Чарльз Иммануэль Форсайт Мейджор на основе двух костей таза описал "гигантскую субфоссильную крысу" Myoryctes rapeto. В 1915 г. ее родовое наименование было заменено на Majoria, поскольку название Myoryctes оказалось уже занятым нематодным червем. Однако, как выяснилось в дальнейшем, тазовые кости Majoria идентичны принадлежащим Plesiorycteropus. В 1912 г. Гийом Грандидье приписал хорошо сохранившуюся бедренную кость крупному родственнику ныне живущего мадагаскарского гигантского хомяка (Hypogeomys antimena), которого он назвал Hypogeomys boulei. Впоследствии эта бедренная кость тоже была отнесена к Plesiorycteropus. Остатки, описанные в качестве Majoria rapeto и Hypogeomys boulei, попадают в верхнюю часть размерного диапазона рассматриваемого рода, что указывает на их вероятную принадлежность Plesiorycteropus madagascariensis. Кроме того, часть таза плезиориктерупуса была приписана вымершей гигантской руконожке (Daubentonia robusta), а в другом случае материал был идентифицирован как принадлежащий крысиному лемуру (Cheirogaleus sp.).

Прорисовки правых тазовых костей: 1 — плезиориктеропус (Plesiorycteropus madagascariensis, = Myoryctes rapeto) (образец M 7085); 2 — водяная полевка (Arvicola amphibius).
В 1946 г. Чарльз Ламбертон на основании большей выбоки уточнил родовой диагноз и отметил существенные различия наличного материала, но не попытался разделить его на несколько видов. Этот ученый отверг утверждение о принадлежности Plesiorycteropus к семейству Orycteropodidae и склонялся к мнению, что он является примитивным, изолированным представителем отряда неполнозубых (Edentata), в который тогда объединялись ксенартры (Xenarthra), трубкозубые (Tubulidentata) и панголины (Pholidota).
В 1975 г. Брайан Паттерсон провел подробную ревизию всех известных на то время остатков и ввиду отличий от других трубкозубых (подсемейство Orycteropodinae) поместил Plesiorycteropus в собственное подсемейство Plesiorycteropodinae, представляющее собой раннее ответвление отряда. Некоторые его морфологические черты, общие с панголинами и другими неполнозубыми, автор счел конвергентными. Также Паттерсон высказал предположение, что предок зверя прибыл на Мадагаскар из Африки в эоцене, примерно в то же время, что и лемуры. В 1985 г. Йоханнес Тевиссен предоставил новые анатомические данные и без комментариев согласился с отнесением Plesiorycteropus к трубкозубым, хотя и раскритиковал некоторые аспекты их классификации Паттерсона. В 2004 г. Инес Хоровиц в морфологическом исследовании, сосредоточенном на южноамериканских копытных (Meridiungulata), поместила Plesiorycteropus среди трубкозубых ближе к их ископаемому роду Myorycteropus, чем к Orycteropus.
В 1994 г. Росс Д.Э. Макфи по частичному черепу из местонахождения Ампасамбазимба в центральной части острова установил вид Plesiorycteropus germainepetterae, отличавшийся от типового меньшими размерами и рядом особенностей строения. Видовой эпитет был присвоен в честь исследователя Жермена Петтера. В той же работе автор с использованием кладистических методов провел исчерпывающий морфологический анализ всего имеющегося материала по Plesiorycteropus. Полученные результаты не дают основания полагать, что данный род в эволюционном отношении относительно ближе к трубкозубым, чем к каким-либо другим копытным (Ungulata) или даже насекомоядным (Insectivora) в широком смысле. Многие черты его скелетного строения, общие с представителями отряда Tubulidentata, могут быть обусловлены функциональной конвергенцией и филогенетической близостью.

Устаревшая реконструкция внешнего облика плезиориктеропуса (Plesiorycteropus madagascariensis). Автор — Х. Шульце.
Чтобы подчеркнуть неопределенное филогенетическое положение Plesiorycteropus среди плацентарных млекопитающих, Макфи учредил монотипический отряд бибималагаший (Bibymalagasia) (bibymalagasy в дословном переводе означает просто "малагасийское животное"). Этот автор полагал, что было бы неприемлемо оставлять современного эвтерия не отнесенным ни к одному отряду, и что открытие большего количества материала или дальнейший анализ вряд ли продемонстрируют близкое родство Plesiorycteropus с любым другим отрядом. Он также считал возможным, но маловероятным, что несколько ископаемых таксонов, таких как Palaeorycteropus и Leptomanis из палеогена Франции, в конечном итоге будут признаны бибималагашиями.
С правомерностью выделения нового отряда согласились многие исследователи (Эшер с соавт., 2003 г.; Зак с соавт., 2005 г.; Холройд и Массер, 2005 г.; Роуз с соавт., 2005 г.; Леманн, 2009 г.). После выдвижения в 1998 г. Станхопом с соавторами концепции клады афротериев (Afrotheria) в нее помимо отрядов Afrosoricida, Macroscelidea, Tubulidentata, Hyracoidea, Proboscidea, Sirenia зачастую стали включать и Bibymalagasia (Хельген, 2003 г.; Эшер, Новачек и Гейслер, 2003 г.).
В 2013 г. Майкл Бакли опубликовал результаты расшифровки последовательностей белка коллагена, выделенного из костей Plesiorycteropus. Полученные данные указывают на наиболее тесную связь последнего с семейством тенрековых (Tenrecidae) — насекомоядными афротериевыми млекопитающими, эндемичными для Мадагаскара. Считается, что тенрековые произошли от общего предка, прибывшего из Африки на плавнике в период между 37 и 29 млн. лет назад. Согласно анализу, Plesiorycteropus и 2 изученных представителя Tenrecidae образуют монофилетическую группу внутри более крупной клады, по отношению к которой златокроты (Chrysochloridae) являются сестринской группой. Бакли предположил, что Plesiorycteropus следует поместить в отряд Tenrecoidea вместе с тенрековыми (Tenrecidae), выдроземлеройковыми (Potamogalidae) и златокротовыми (Chrysochloridae); последние два семейства отделились от первого около 53-47 и 69-59 млн. лет назад соответственно. За рамками молекулярно-генетического исследования осталось большинство родов тенреков и выдровые землеройки, что оставляет открытой возможность отнсения Plesiorycteropus к подсемейству Plesiorycteropodinae в пределах семейства Tenrecidae, либо выделения в семейство Plesiorycteropodidae.

Компьютерные модели частичных черепов плезиориктеропусов c лабиринтом внутреннего уха: А — Plesiorycteropus germainepetterae (типовой образец MNHNР 327); В — Plesiorycteropus madagascariensis (типовой образец MNHNР 328).
Сравнительное исследование структуры костного лабиринта внутреннего уха, проведенное Бенуа с коллегами в 2015 г., подчеркнуло филогенетическую близость Plesiorycteropus, с одной стороны, с кладой Afrosoricida + Macroscelidea и, с другой стороны, с Tubulidentata, а также определил, что Plesiorycteropus madagascariensis и Plesiorycteropus germainepetterae могут не являться отдельными видами.
Строение и функции
Остатки плезиориктеропусов достаточно фрагментарны, так что данные о строении их скелета также весьма неполные. Найдены четыре частичных черепа (три из них принадлежат Plesiorycteropus madagascariensis и еще один — Plesiorycteropus germainepetterae), некоторые позвонки, элементы плечевого и тазового поясов, а также разрозненные кости конечностей целого ряда особей.
Общие размеры плезиориктеропусов определить трудно. Длина лобной кости Plesiorycteropus madagascariensis составляет в среднем 35,4 мм, Plesiorycteropus germainepetterae — 29,4 мм. Грубые расчеты предполагают, что Plesiorycteropus madagascariensis имел череп длиной приблизительно в 10 см, совокупную длину головы и туловища около 60-80 см и общую длину более 1 м. Масса тела, рассчитанная по площади поперечного сечения бедренной кости на основе сравнительных данных по броненосцам, панголинам, свинкообразным грызунам и приматам, составляет 9-18 кг для Plesiorycteropus madagascariensis и 6-10 кг для Plesiorycteropus germainepetterae. Низкие оценки выглядят более предпочтительно, поскольку основаны на сравнении с бедренными костями броненосцев, сходными с таковыми плезиориктеропусов. Однако по любой из оценок эти животные значительно превосходят крупнейшее ныне живущее тенрекообразное — обыкновенного тенрека (Tenrec ecaudatus) весом до 2 кг.

Частичный череп плезиориктеропуса (Plesiorycteropus madagascariensis) (образец MNHNР 534) из местонахождения Амбулисатра, Мадагаскар; снизу.
У трех известных черепов отсутствует лицевой отдел, причем он отломлен примерно в одном и том же месте — в околоносовых полостях, в передней части мозговой коробки. Это указывает на то, что передняя часть черепа была более тонкой и хрупкой, чем задняя часть, состоящая из толстых костей. Судя по сохранившимся у четвертого черепа носовым костям, морда была достаточно длинной и по своим очертаниям напоминала таковую броненосцевых (Dasypodidae). Крепкие носовые кости уплощены. Они расширяются кпереди, что довольно необычно для плацентарных и также наблюдается у броненосцев. Носовая перегородка, разделяющая левую и правую носовые полости, окостенела в большей степени, чем у большинства других млекопитающих; подобное состояние отмечено только у ленивцев, у которых очень короткий нос.
Мозговой отдел черепа вытянут. Слезная кость относительно большая. У нее есть один слезный канал, который открывается около шва между лобной и слезной костями, как у эвлипотифлей. Рядом с этим отверстием имеется небольшой бугорок, отсутствующий у трубкозубов. Глазничная полость, в которой находится глазное яблоко, относительно короткая, что напоминает ситуацию у панголинов и броненосцев. В кости глазницы имеется круглое отверстие. Как у ксенартров и панголинов, сверху видна лишь небольшая часть чешуйчатой кости. На шве между лобной и теменной костями имеется височный бугорок. Подобно трубкозубам, теменные кости относительно большие. Присутствует межтеменная кость. В отличие от муравьедов и панголинов, затылок плоский и вертикально ориентированный. Нет выемок над большим затылочным отверстием, которые имеются у трубкозубов. Каменистая кость образует относительно большую часть крыши барабанной полости, в которой находится среднее ухо.
Ямка снизу черепа для причленения нижней челюсти мелкая, что предполагает слабость челюстной мускулатуры. В задней части этой ямки находится псевдогленоидный отросток. Строение нижней челюсти остается неизвестным, но по всей вероятности она не отличалась массивностью. О наличии зубов также ничего неизвестно; не исключено, что в связи с мирмекофагией они были полностью утрачены. Судя по эндокранам, полушария переднего мозга были небольшими. Малая ширина канала зрительного нерва подразумевает небольшой размер глаз и слабое зрение, как у многих других тенрекообразных. Наоборот, значительные по объему носовые полости с сильно развитыми обонятельными раковинами свидетельствуют о хорошем обонянии, что также характерно для данного подотряда.
Позвоночный столб включает 5 или 6 поясничных и как минимум 7 крестцовых позвонков. Нет никаких свидетельств наличия дополнительных суставов между позвонками, которые характерны для ксенартров. У известных грудных позвонков сочленения с межпозвоночными дисками имеют фасолевидную форму и намного шире в поперечном направлении, чем сверху вниз. Лопатка, отнесенная к Plesiorycteropus лишь условно, обладает акромиальным отростком, но, вероятно, не настолько большим, как у трубкозубов или броненосцев. Таз составлен парой узких подвздошных и парой длинных седалищных костей.
Конечности сильные, задние длиннее передних. Плечевая кость мощная, с энтепикондилярным отверстием на дистальном конце. Лучевая кость компактная и массивная, сходная с таковой панголинов. Локтевая кость с хорошо развитым локтевым отростком на проксимальном конце, тогда как ее дистальный конец сужен; общее строение кости предполагает, что передняя конечность могла развивать значительное усилие. Бедренная кость с длинной шейкой, как у обыкновенной гимнуры (Echinosorex gymnura), и с III-м вертлугом. Большая и малая берцовые кости широко сращены в тибиофибулу. Последняя напоминает эту структуру у броненосцев обширным сращением, сжатием стержня большой берцовой кости, узостью сочленовной поверхности на дистальном конце и широким пространством между обеими костями. В отличие от броненосцев, большая и малая берцовые кости не наклонены друг относительно друга, а располодены почти параллельно. Таранная кость широкая и короткая, с исключительно большим заднемедиальным отростком. Пястные и плюсневые кости достаточно короткие, широкие проксимально и узкие дистально. Проксимальные фаланги короче средних, а дистальные фаланги узкие и когтеобразные.

Правая плечевая кость плезиориктеропуса (Plesiorycteropus sp.) (образец MNHNA 1987.036).
Передняя конечность плезиориктеропусов демонстрирует комплекс копательных адаптаций: лучевая кость уступает по длине плечевой, пястные кости короткие и широкие, а конечные фаланги пальцев поперечно сужены, заострены и могли быть вооружены крупными когтями. Передняя лапа помещалась на субстрат, когти входили в него, а затем лапа оттягивалась назад к телу, взрывая грунт. Ряд признаков также указывает на ограниченную способность к лазанию по деревьям. Помимо вероятности наличия больших когтей, общее строение передней конечности напоминает таковое у землеройковых тенреков (Microgale spp.) и гимнур (Galericinae), которые могут лазать и тоже имеют маленькие глаза. Головка плечевой кости возвышается над окружающей бугристостью, облегчая поднятие передней конечности до уровня плеча или выше.
Некоторые особенности строения позвоночника и тазовых костей предполагают, что плезиориктеропусы часто принимали вертикальное или сидячее положение. В частности, седалищные бугры таза приобрели вид широких плато треугольной формы с гладкой поверхностью, в отличие от узких угловатых выступов большинства плацентарных. Сходные бугры имеются у мартышковых обезьян (Cercopithecidae) ислужат основанием для седалищных мозолей. Хвост достаточно мощный, особенно толстый у основания. Находка серии проксимальных позвонков хвоста, медленно уменьшающихся в размере спереди назад, предполагает, что он был длинным.
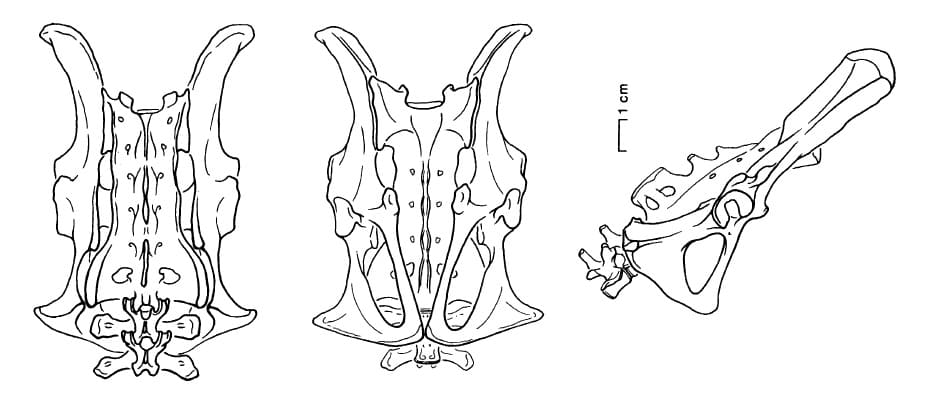
Прорисовки реконструированного таза плезиориктеропуса (Plesiorycteropus madagascariensis).
От всех других известных эвтериев рассматриваемый род отличает следующая комбинация производных признаков: носовые кости заметно расширены кпереди (носовые кости Orycteropus узкие спереди и расширяются в заднем направлении); нижнечелюстная ямка широкая и плоская, ограниченная сочленовной фацией без вовлечения скулового отростка чешуйчатой кости (у Orycteropus эта ямка умеренно глубокая и также не затрагивает скуловой отросток); невральные дуги задних грудных и всех поясничных позвонков пронизаны крупными продольными каналами (у Orycteropus имеются лишь крошечные и не перфорирующие зачатки таких каналов); на тазовых костях седалищные бугры сильно расширены и уплощены сзади (у Orycteropus такое расширение отсутствует); на таранной кости имеется очень большой заднемедиальный отросток с глубокой бороздкой на нижней стороне, вероятно, для прохождения сухожилия мышцы-сгибателя (таранная кость Orycteropus также имеет заднемедиальный отросток, но у него отсутствует отчетливая нижняя бороздка).
Состав
Plesiorycteropus madagascariensis Filhol, 1895 (тип) — типовой вид, известный из ? позднего плейстоцена - голоцена северо-западной, центральной и юго-западной частей Мадагаскара (местонахождения Амбулисатра, Ампасамбазимба, Анджохибе, Антсирабе, Бело-сур-мер). Синонимы: Myoryctes rapeto Forsyth Major, 1908; Hypogeomys boulei Grandidier, 1912; Majoria rapeto (Forsyth Major, 1908) Thomas, 1915.
Описан по черепу без лицевого отдела, по-видимому, принадлежащему почти взрослой особи.

Частичный череп плезиориктеропуса (Plesiorycteropus madagascariensis) (типовой образец MNHNР 328) из местонахождения Бело-сур-мер, Мадагаскар; сбоку, сверху, снизу и сзади.
По сравнению с Plesiorycteropus germainepetterae, общие размеры больше; мозговая коробка больше и не столь шаровидна; межглазничное сужение менее выражено; височные линии расположены выше; псевдогленоидный отросток менее выражен; рядом с овальным отверстием нет небольшого отверстия; передние и задние барабанные отростки каменистой кости более развиты; височный бугорок слабо выражен; верхний профиль затылочного гребня прямой; III-й вертлуг бедренной кости больше.
Plesiorycteropus germainepetterae MacPhee, 1994 — вид, известный из ? позднего плейстоцена - голоцена центральной части Мадагаскара (местонахождение Ампасамбазимба); ему также могут принадлежать некоторые кости из ряда местонахождений юго-западной части острова.
Описан по черепу без лицевого отдела, принадлежащему взрослой особи.

Частичный череп плезиориктеропуса (Plesiorycteropus germainepetterae) (типовой образец MNHNР 327) из местонахождения Ампасамбазимба, Мадагаскар; сбоку, сверху и снизу.
По сравнению с Plesiorycteropus madagascariensis, общие размеры меньше (линейно на 8-19 %); мозговая коробка меньше и более шаровидная; межглазничное сужение выражено сильнее; височные линии расположены ниже; псевдогленоидный отросток более выражен; рядом с овальным отверстием имеется небольшое, вероятно, сосудистое отверстие; передние и задние барабанные отростки каменистой кости менее развиты; височный бугорок выступающий; верхний профиль затылочного гребня вдавлен посередине; III-й вертлуг бедренной кости меньше. По мнению ряда исследователей, самостоятельность данного таксона спорна, а его особенности могут объясняться половым диморфизмом и индивидуальной изменчивостью.
Остатки, определенные лишь до рода и обозначенные Plesiorycteropus spp., известны из голоцена центральной и юго-западной частей Мадагаскара (местонахождения Амбулисатра, Ампасамбазимба, Ампоза, Анджохибе, Ановаха, Антсирабе, Бемифани, Масинандраина, Сираве и, возможно, Анкево).

Карта местонахождений остатков Plesiorycteropus: зеленый цвет — Plesiorycteropus madagascariensis, синий цвет — Plesiorycteropus madagascariensis и Plesiorycteropus germainepetterae, красный цвет — Plesiorycteropus spp.
Образ жизни
Субфоссильные остатки плезиориктеропусов происходят из дюжины местонахождений Мадагаскара, расположенных как в лесистых районах вдоль его побережий, так и в открытых местностях внутри острова. В последних областях эти звери могли населять лесные заросли, окружающие большие заболоченные водоемы и в дальнейшем исчезнувшие. Не исключено, что ареалы Plesiorycteropus madagascariensis и Plesiorycteropus germainepetterae довольно широко перекрывались, поскольку остатки этих видов выявлены по меньшей мере в одном общем местонахождении.
По-видимому, плезиориктеропусы являлись довольно громоздкими животными, живущими на поверхности земли, однако не исключена их способность забираться на деревья. Полагаясь на острый нюх, они в темное время суток охотились на муравьев и термитов, но при случае потребляли и другую мягкую пищу, такую как дождевые черви и личинки жуков. В поисках корма эти звери копались в лесной подстилке или сдирали кору с гнилых стволов когтистыми передними лапами, при необходимости опираясь на задние конечности и хвост, либо присаживаясь на гузно. Можно предположить, что плезиориктеропусы вскрывали гнезда колониальных насекомых, но ввиду относительно небольших размеров они вряд ли могли вламываться в затвердевшие термитники, как это делает современный трубкозуб. Укрывались животные в норах, между толстых корней деревьев, под нагромождениями валежника и в других подобных местах.

Плезиориктеропус (Plesiorycteropus madagascariensis) в естественном окружении. Автор — Э. Степп.
Вымирание
Обстоятельства вымирания рода доподлинно неизвестны. По всей видимости, оно приурочено к периоду между I и X вв., когда исчез целый ряд представителей мадагаскарской фауны. Очевидно, этому способствовала хозяйственная деятельность человека, включая охоту и выжигание лесов. При этом вымирание плезиориктеропусов достаточно уникально в том отношении, что многие другие исчезнувшие виды (например, крупные лемуры и фосса, небольшие бегемоты, гигантские нелетающие птицы) были гораздо больше по размерам и не являлись насекомоядными, тогда как некоторые животные с более специфичным рационом (в частности, мелкозубый мунго (Eupleres goudotii) и руконожка (Daubentonia madagascariensis)) выжили.
Находки остатков плезиориктеропусов в ассоциации с костями и следами деятельности человека неизвестны. Фрагмент таза из Масинандраины с помощью радиоуглеродного метода была датирован II в. до н. э., то есть временем вскоре после прибытия на Мадагаскар австронезийских поселенцев. В XVII в. вропейские исследователи уже не обнаружили на острове ничего похожего на плезиориктеропусов, а в местном фольклоре о них не сохранилось никаких упоминаний. Впрочем, радиоуглеродное датирование костных остатков другого млекопитающего, найденных вместе с костями Plesiorycteropus madagascariensis в пещерах Анджохибе на северо-западе Мадагаскара, позволяет предположить, что он продолжал существовать менее 500 лет назад.
Ссылки
Plesiorycteropus (Paleobiology Database) перейти
Plesiorycteropus (Wikipedia) перейти
Литература
Каландадзе Н. Н., Шаповалов А. В. Судьба мегафауны наземных экосистем Мадагаскарской зоогеографической области в позднем антропогене.
Мадагаскар / Науч. ред. Б. Д. Васильев. — М.: Прогресс, 1990
Россолимо О. Л., Павлинов И. Я., Крускоп С. В., Лисовский А. А., Спасская Н. Н., Борисенко А. В., Панютина А. А. Разнообразие млекопитающих. Ч. 1-3. — М.: Изд-во КМК, 2004
Cenozoic Mammals of Africa. Edited by Werdelin L. & Sanders W. J. — Berkeley: University of California Press, 2010
Nowak R. M. Walker’s Mammals of the World. Monotremes, Marsupials, Afrotherians, Xenarthrans, and Sundatherians. — Baltimore: Johns Hopkins University Press, 2018