Проайлуры — Proailurus
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Териеобразные (Theriiformes)
Клада Голотерии (Holotheria)
Клада Трехнотерии (Trechnotheria)
Клада Кладотерии (Cladotheria)
Клада Затерии (Zatheria)
Клада Трибосфениды (Tribosphenida)
Клада Терии (Theria)
Клада Эвтерии (Eutheria)
Клада Плацентарные (Placentalia)
Магнотряд Бореоэвтерии (Boreoeutheria)
Клада Скротиферы (Scrotifera)
Клада Феревунгуляты (Fereuungulata)
Клада Ферэ (Ferae)
Клада Всехищные (Pan-Carnivora)
Клада Хищнообразные (Carnivoramorpha)
Отряд Хищные (Carnivora)
Подотряд Кошкообразные (Feliformia)
Клада Элуроиды (Aeluroidea)
Надсемейство Фелоиды (Feloidea)
Семейство Кошачьи (Felidae)
Подсемейство †Проайлурины (Proailurinae)
Род †Проайлуры (Proailurus)
Вид †Proailurus lemanensis
Вид †Proailurus bourbonnensis
Вид †Proailurus major
Проайлуры (Proailurus Filhol, 1879) — род монотипического подсемейства проайлурин (Proailurinae Zittel, 1893), происходящий из позднего олигоцена - раннего миоцена Европы. Установлены 3 вида.
Реконструкция внешнего облика проайлура (Proailurus lemanensis). Автор — Р. Евсеев.
Синоним: Proaelurus Forbes, 1881.
История изучения
Род Proailurus установил французский натуралист Генри Фильоль в 1879 г. по остаткам, найденным в коммуне Сен-Жеран-ле-Пюи на северо-западе Франции. Родовое нраименование образовано древнегреческими словами πρό- (pro-) — "перед, прешествующий" и αἴλουρος (ailurus) — "кошка". Первоописатель выделил два вида — Proailurus lemanensis и Proailurus julieni. Однако второй вид позже был помещен в род Stenogale. В 1882 г. Фильоль описал третий вид, Proailurus medius. В 1888 г. М. Шлоссер принял его в качестве типового представителя рода Haplogale. В 1998 г. Р. Хант сравнил черепа ряда элуроидов и подтвердил валидность нового рода. В 1999 г. С. Пейн провел ревизию рода, в ходе которой выделил еще два вида — Proailurus boubonnensis и Proailurus major.
Строение и функции
Размеры тела типового вида Proailurus средние для кошек: длина его черепа составляет около 15 см, общая длина головы и туловища — свыше 60 см, высотой в плечах — порядка 38 см, масса тела — около 9 кг. Примерно такой же величины достигает современная рыжая рысь (Lynx rufus). Расчетный вес других представителей рода варьирует в пределах 7-23 кг. Эти коренастые животные имели вытянутое туловище, относительно короткие конечности и длинный хвост, напоминая телосложением одновременно кошек и виверр.
Череп в лицевом отделе удлинен сильнее, чем у современных кошек, что усиливает его сходство с черепом виверр. Заглазничное сужение напоминает таковое других кошкообразных хищников. Сагиттальный гребень высокий и тонкий. Ярко выраженный затылочный гребень образует срединный выступ, выступающий назад за уровень затылочных мыщелков. Основная клиновидная и основная зытылочная кости широкие. Основная крылоклиновидная кость смещена назад сильнее, чем у ранних нимравид, а ее овальное отверстие занимает переднее положение, как и у современных кошачьих. Строение слуховой области во многом аналогично таковому ныне живущих кошек, демонстрируя вполне современный тип слухового барабана, однако, в слуховом регионе Proailurus имеется хорошо развитый нижний отросток мыска верхнего края пирамиды височной кости, отсутствующий у современных кошачьих.
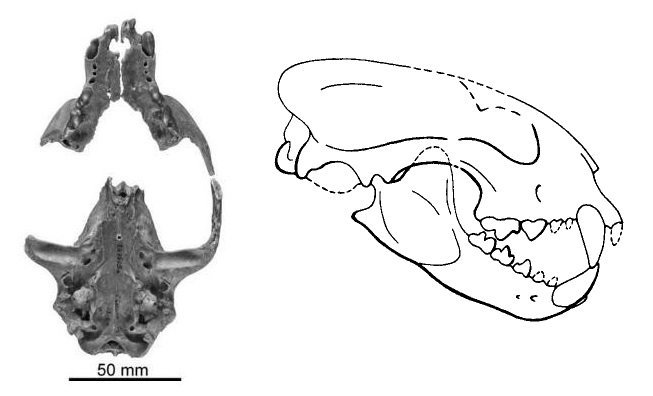
Вверху и внизу слева — типовой неполный череп с нижней челюстью проайлура (Proailurus lemanensis), внизу справа — прорисовка его частичной реконструкции.
Массетерная ямка глубокая. Венечный отросток короткий и ориентирован вертикально. Он все еще относительно вертикален у более молодых североамериканских таксонов Pseudaelurus validus и Pseudaelurus skinneri, но тонкий и скошенный у Pseudaelurus intrepidus и современных кошачьих. По размерам венечный отросток меньше, чем у Pseudaelurus и признаков характерного "крюка", как у современных кошек, нет. В целом, по черепному строению Proailurus весьма напоминает Pseudaelurus, но череп последнего имеет гораздо большую величину и, соответственно, более крупные челюсти.
P3 широкий, но менее массивный, чем у Pseudaelurus. Он имеет парастиль на передней поверхности, не отличаясь этим признаком от Pseudaelurus или современных кошачьих. Его крупный протокон расположен передне-лингвально. Протокон P4 расположен ближе к внутренней стороне и находится под немного более тупым углом к лингвальной поверхности зуба. На задней поверхности основных выступов P3 и P4 имеются дополнительные бугорки, отделенные четко выраженным пояском. Эта деталь зубной морфологии также наличествует у Pseudaelurus и сохраняется с небольшим изменением у современных кошачьих.
Восстановленный череп с нижней челюстью проайлура (Proailurus lemanensis) (образец AMNH 101931). Американский музей естествознания, Нью-Йорк.
M1 сжат в продольном направлении, но расширен поперек. Паракон и метакон (всегда присутствующий у Proailurus) почти слиты, лингвальный край образован большим протоконом — все эти признаки характерны и для Pseudaelurus, резко контрастируя с рудиментарным M1 современных кошек.
Функциональность хищнических зубов выглядит относительно примитивной — паракон M1 граничит с лезвием метастиля P4, таким образом удлиняя режущую поверхность. Оба эти зуба отличаются от соответствующих у современных кошачьих не только своим относительно большим размером и меньшим поперечным диаметром, но также и отсутствием заднего пояска. У Proailurus эти зубы очень походят на таковые мадагаскарского хищника Сryptoprocta.
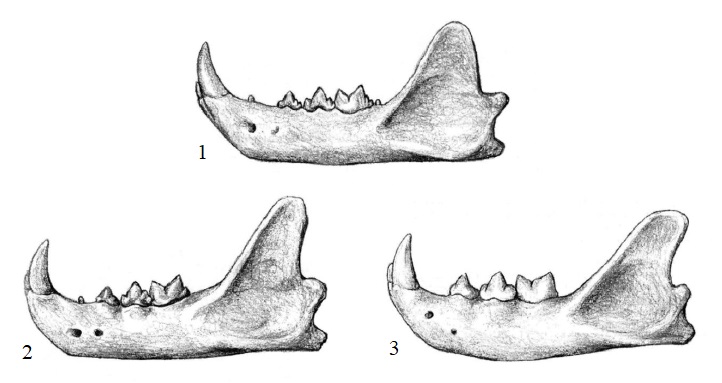
Прорисовки нижних челюстей Proailurus (1), Pseudaelurus (2) и современной кошки (3). Автор — М. Антон.
M1 демонстрирует очевидные признаки происхождения от зубов бугорчато-режущего типа. Передний выступ сформирован так же, как у кошачьих, но задний бугор выше и менее сглажен, обладая отчетливыми углами между боковыми и задними поверхностями. На многих образцах присутствует небольшой, но хорошо различимый бугорчато-режущий выступ, завершающий примитивный треугольник бугорчато-режущего коренного зуба.
Зубная морфология Proailurus в оцелом сходна с таковой более продвинутых кошек, но сохраняет некоторые элементы, полностью редуцированные у современных форм. Так, с каждой стороны нижней челюсти имеется 3 предкоренных и 2 коренных зуба, тогда как у Pseudaelurus их уже 3 и 1 соответственно, а диастема между нижним клыком и р3 начала сокращаться. От современных кошек Proailurus отличается варьирующим наличием Р1, p1, p2 и m2, а также присутствием небольшого метаконида и талонида на m1. Эти признаки считаются примитивными в связи с тем, что во многих эволюционных линиях некоторые зубы, как правило, становятся специализированными, в то время как другие редуцируются или даже исчезают.
Череп с нижней челюстью и портретная реконструкция проайлура (Proailurus lemanensis). Автор — Р. Евсеев.
Позвоночный столб остается в большей степени неизвестным, однако не подлежит сомнению, что он был столь же длинным и гибким, как у более позднего Pseudaelurus и современных кошачьих.
Хотя значительная часть скелета Proailurus пока не обнаружена, врядли он кардинально отличался от такового Pseudaelurus. Если Proailurus, Pseudaelurus и Lynx имеют приблизительно одинаковую высоту в плечевой и тазовой областях, то Lynx демонстрирует несколько более удлиненные пястные и плюсневые кости и несколько более короткую плечевую кость, чем обе миоценовые кошки.
Плечевая кость схожа с таковой современных кошек, но ключичная и плечевая впадины плечевого сустава имеют большую площадь и более грубую поверхность срединных надмыщелков в отличие от небольших и гладких надмыщелков современных представителей семейства. Эта комбинация увеличенного срединного надмыщелка и глубокой ямки локтевого отростка предполагает, что
Proailurus был умеренно курсориален. Ближняя головка локтевой кости имеет овальную форму. Кость сужена в области шейки, как и у современных Felidae и расширяется к нижнему концу, как у
Pseudaelurus.
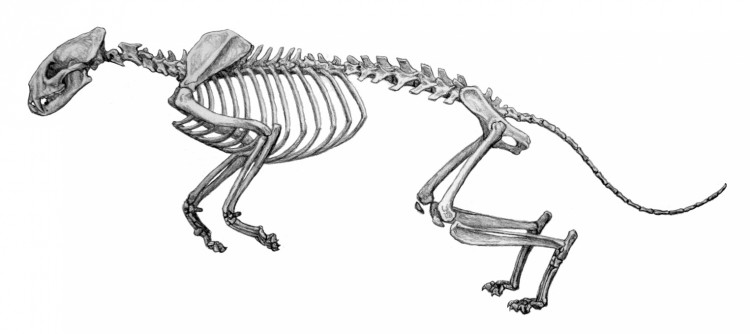
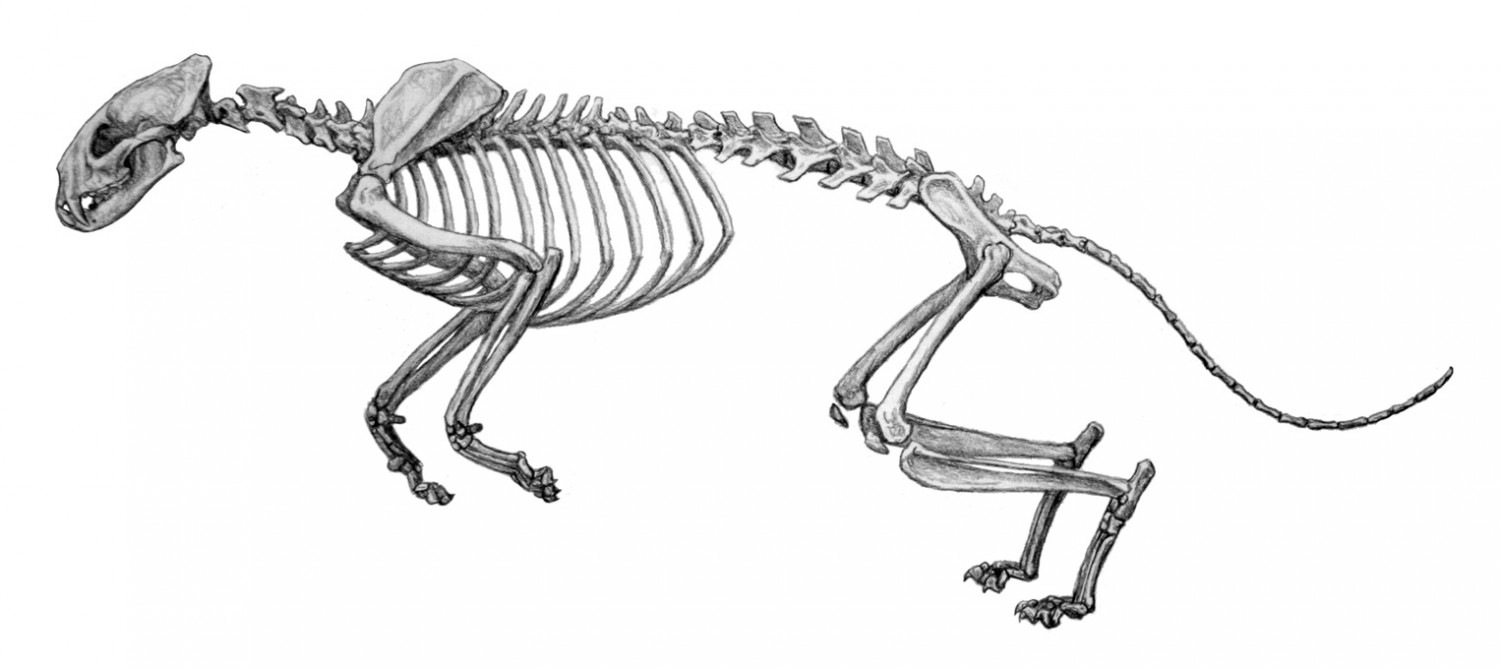
Прорисовка скелета проайлура (
Proailurus lemanensis), реконструированного с дополнением остатков
Pseudaelurus. Автор — М. Антон.
Пястные кости схожи с таковыми у Pseudaelurus. I-я пястная кость имеет тонкое тело и параллельную земле дальнюю суставную поверхность. Данные о пястных костях и пальцах ступни указывают, что при небольшом изменении в пропорциях этих элементов в пределах семейства Felidae, начиная с раннего миоцена, плюсневые кости особых изменений не претерпели.
В целом, в отношении передних конечностей наблюдается тенденция к пропорциональному изменению размеров плечевой, локтевой и пястных костей — если соотношение их длины Proailurus и Pseudaelurus практически идентичны, то у Felis и Panthera плечевые кости укорачиваются, занимая примерно 40 % от общей длины конечности, в отличие от 45 % у Proailurus. В соотношении длины костей кисти к длине конечностей наблюдается обратная картина — 12 % у Proailurus и 15 % у Felis и Panthera.

Реконструкция внешнего облика проайлура (
Proailurus lemanensis). Автор — М. Антон.
Бедро длинное и тонкое, напоминая таковое многих архаичных хищных (Dinictis, Cynodictis, Cryptoprocta). Одной из особенностей является наличие хорошо развитого III-го вертлуга, представляющего собой продолжение внешней шероховатой линии, что отмечено также у Amphicyon, Cynodictis и Dinictis. Голень значительно короче, чем бедро и при взгляде спереди не имеет бокового искривления, которое присутствует у многих примитивных элуроидов. Углубление для таранной кости сравнительно большое.
Пяточный бугор короткий и толстый, напоминая таковой у Dinictis. Задний выступ таранной кости большой, загнутый назад. Внешняя сторона дальнего конца пяточной кости имеет большой выступ, который хорошо развит у стопоходящих животных, однако в то же время присутствует у таких форм, как тигр. I-я плюсневая кость рудиментарна, как у других кошачьих. По сравнению с Proailurus, соотношение отдельных сегментов задней конечности современных кошачьих не претерпело особых изменений.
Состав
Proailurus lemanensis Filhol, 1879 (тип) — типовой вид, известный из позднего олигоцена (MP 30, хаттский ярус) - раннего миоцена (MN 2b, аквитанский ярус) Франции (регионы Овернь-Рона-Альпы и Окситания). Описан по неполному черепу с нижней челюстью (образцы MNHN-SG 3509a-e). Помимо этого, имеется другой зубочелюстной материал. Вес животного оценивается приблизительно в 9 кг.
Типовой неполный череп с нижней челюстью проайлура (Proailurus lemanensis).
Proailurus bourbonnensis Peigne, 1999 — вид, известный из раннего миоцена (MN 2a, аквитанский ярус) Франции (регион Овернь-Рона-Альпы) и Германии (земля Рейнланд-Пфальц). Синонимы: Proailurus lemanensis Filhol, 1879 (частью); Proailurus sp. Ginsburg, 1983. Описан по левому нижнечелюстному фрагмениту с p3-m2 (образец MNHN-Lim 593). Также найдены другие фрагменты н ижних и верхних челюстей и разрозненные зубы. Расчетный вес животногосоставляет 7-10 кг. Из зубов p1 полностью отсутствует, m1 несколько менее удлинен, а m2 в меньшей степени редуцирован, чем у Proailurus lemanensis.

Прорисовки типового левого нижнечелюстного фрагмента проайлура (Proailurus bourbonnensis).
Proailurus major Peigne, 1999 — вид, известный из раннего миоцена (MN 2a, аквитанский ярус) Франции (регион Овернь-Рона-Альпы). Описан по левому нижнечелюстному фрагменту с p3-m1 (образец MHNT-GER 263). Размеры наибольшие в роду: по оценкам, вес мог достигать 23 кг. Зуб p4 относительно крупнее, а m1 менее удлинен, чем у Proailurus lemanensis.

Прорисовки типового левого нижнечелюстного фрагмента проайлура (Proailurus major).
Остатки, определенные лишь до рода и обозначенные Proailurus sp., известны из раннего миоцена (MN 2, аквитанский ярус) Испании (автономное общество Арагон).
За пределами Европы достоверные остатки Proailurus не обнаружены. Р. Хант (1998 г.) обозначил в качестве Proailurus sp. нижнечелюстной фрагмент из раннеолигоценовых отложений формации Хсанда-Гол в Монголии. Однако С. Пэйн (1999 г.) пришел к заключению, что данный экземпляр представляет собой нимравида из рода Eofelis, с чем согласились Л. Верделин с коллегами (2010 г.). Т. Ротуэлл (2004 г.) указал на его принадлежность к виду кошачьих Pseudaelurus cuspidatus.
Кроме того, Хант (1998 г.) описал целый череп Proailurus из раннемиоценовых (верхи хемингфордского яруса) слоев местонахождения Гинн-Куорри в штате Небраска, США. Данный образец составляет 14,2 см длину, отличается массивностью и обладает крупными и толстыми клыками. Этот автор также отметил ряд экземпляров, которые, по его мнению, могут относиться к Proailurus, включая несколько из формации Шип-Крик в штате Небраска: один величиной с рысь, другой с небольшого леопарда. Кроме того, Хант упоминает экземпляр из Ист-Куямунге в штате Нью-Мексико, возможно, того же вида, что и более крупная особь из Шип-Крик; и еще один экземпляр из Эхо-Куорри. Однако более крупный экземпляр из Шип-Крик, а также экземпляры из Куямунг и Эхо-Куорри были отнесены к виду Pseudaelurus validus в 2001 г., а меньший экземпляр из Шип-Крик — к Pseudaelurus skinneri в 2003 г. Кошка из Гинн-Куорри была описана, но не отнесена ни к одному таксону Г. Лирасом с коллегами в 2019 г., хотя авторы заявили, что ее не следует относить к кошачьим "проайлуринового" класса, и предложили отнести ее к Hyperailurictis (несмотря на то, что она значительно древнее любого известного представителя данного рода) или к новому роду.
Согласно Ханту, основание черепа из Гинн-Куорри по своей структуре весьма плезиоморфично европейскому Proailurus. Это предполагает, что филогенетическое разнообразие Felidae началось раньше миоцена и что североамериканский Pseudaelurus, возможно, развился из предка Proailurus, а не из раннего Pseudaelurus, мигрировавшего в Северную Америку. Если это так, то кошачьи, возможно, распространились проникли на территорию Американского континента уже в начале хемингфордия вместе со многими другими родами хищных — Cynelos, Ysengrinia, Amphicyon, Cephalogale, Phoberocyon, Ursavus, Potamotherium, Edaphocyon (Цю, 2003 г.).
Образ жизни
Проайлуры являлись обитателями закрытых ландшафтов лесного типа, существовавших в условиях субтропического климата. Обладая превосходно развитым зрением, острыми зубами и, по всей видимости, полувтяжными когтями, они успешно охотились на мелких млекопитающих: землероек, ежей, пищух, кроликов, белок и многих других грызунов, оленьков, кайнотериев и различных птиц. Конкуренцию этим хищникам могли составлять, например, мелкие амфициониды, от которых они, возможно, укрывали свою добычу на древесных ветвях.
Проайлур (Proailurus lemanensis) с пойманным грызуном. Автор — Р. Учитель.
Вполне вероятно, что проайлуры были хорошо приспособлены к передвижению по деревьям и добывали здесь часть своей пищи, демонстрируя сходство с такими современными элуроидами, как генетта (Genetta genetta), пальмовая цивета (Nandinia binotata) и фосса (Сryptoprocta ferox). Длинные задние конечности обеспечивали эффективный толчок, гибкий позвоночник обеспечивал маневренность, а короткое плечо и пальцы кисти гарантировали крепкий захват. Вероятно, проайлуры были еще стопоходящими, а не пальцеходящими, как современные кошки. Стопоходящее положение ступни, с большей площадью контакта между ней и опорной поверхностью, является более устойчивым и безопасным при древолазании, чем пальцеходящее. Широкий диапазон совместных движений суставов лодыжки и запястья позволяет ступням лучше приспособиться к неровному субстрату. Все это свидетельствует о том, что проайлуры, вероятно, были гораздо лучшими эквилибристами, чем ныне живущие кошки. В данном отношении они могли больше напоминать фоссу, успешно охотящуюся в кронах деревьев на чрезвычайно проворных сифак и других лемуров.
Ранний миоцен Сен-Жеран-ле-Пюи, Франция. В субтропическом лесу проайлур (Proailurus lemanensis) точит когти о ствол дерева, а с другого дерева за ним наблюдает белка палеосциур (Paleosciurus sp.). Автор — М. Антон.
В ходе дальнейшей эволюции кошки в разной степени отступили от этого состояния, разработав более эффективные формы наземного передвижения. Это видно уже из скелета
Pseudaelurus, демонстрирующего удлинение метаподиев и уменьшение степени свободы в боковом движении голеностопного сустава по сравнению с
Proailurus. Последняя морфологическая черта является особенно важной, поскольку ограничение бокового вращения сустава повышает эффективность движения в вертикальной плоскости и этот способ лучше подходит для передвижения по земле.
На древесных ветвях самка проайлура (Proailurus lemanensis) кормится тушей добытого кайнотерия (Cainotherium sp.), в то время как ее детеныши заняты игрой. Автор — М. Антон.
Род Proailurus был учрежден Фильолем (1879 г.) для обозначения образцов из позднего олигоцена Франции, обладающих многими плезиоморфными признаками Pseudaelurus, но более древних в биостратиграфическом отношении. Коп (1880 г.) первым предположил, что Proailurus и Pseudaelurus являлись одними из первых представителей семейства кошачьих. К концу XIX в. родство олигоцен-миоценового Proailurus, миоценового Pseudaelurus и ныне живущих кошек уже не вызывало сомнения (Адамс, 1897 г.).
Установление родственных связей между различными родами и видами внутри семейства Felidae до сих пор в значительной мере затруднено. Pseudaelurus — наиболее ранний представитель семейства кошачьих в Северной Америке, чей скелет в большей мере сходен с Proailurus, чем с Felis. Временные рамки существования всех трех родов никак не перекрываются. И все же, независимо от того, представляют ли Proailurus и Pseudaelurus монофилетическую группу, оба этих рода считаются самыми ранними и примитивными членами семейства Felidae.
Проайлур (Proailurus lemanensis) в естественном окружении. Авторы — Р. Ломотан (слева) и В. Симеоновски (справа).
По имеющимся палеонтологическим и молекулярным данным, кошкообразные (Feliformia) возникли в конце эоцена (Гобер и Верон, 2003 г.). Наиболее ранние формы, близкие к фелоидным линиям, Stenogale и Haplogale, ведут родословную с эоцен-олигоценовой границы. В связи с этим вполне допустимо предположить, что семейство кошачьих оформилось в раннем олигоцене (Градштайн, 2004 г.). Первой "настоящей" кошкой считается Proailurus, известный начиная с позднего олигоцена.
Таксономическое положение этого древнего хищника и сейчас не совсем ясно: филогенетический анализ (Весли-Хант и Флинн, 2005 г.) показал, что Proailurus является базальным представителем надсемейства Feloidea, таким образом находясь близко к общему предку кошачьих (Felidae), виверровых (Viverridae), мангустовых (Herpestidae) и гиеновых (Hyaenidae). Однако другие исследования (Розуэлл, 2003 г.) подтверждают принадлежность Proailurus непосредственно к семейству Felidae, где для него установлено отдельное подсемейство Proailurinae.
Proailurus считается возможным предком родовой группы "Pseudaelurus", существовавшей в миоцене и, вероятно, давшей начало основным линиям кошачьих — махайродонтинам (Machairodontinae) и фелинам (Felinae), хотя филогенетическое положение примитивных представителей семейства до сих пор остается не совсем ясным. Несмотря на ряд особенностей, общее строение Proailurus было уже явно кошачьим, а к середине миоцена такие роды как Styriofelis достигли почти современных морфологических адаптаций.
Ссылки
Antón M. The beginnings of catkind:
Proailurus. — 2014 (Chasing sabretooths)
перейтиProailurus (Paleobiology Database)
перейти
Литература
Тернер А., Антон М. Большая энциклопедия доисторических животных. — М.: Оникс, 2006
Augusti J., Antón M. Mammoths, sabertooths, and hominids: 65 million years of mammalian evolution in Europe. — New York: Columbia University Press, 2002
Rose K. D. The Beginning of the Age of Mammals. — Oxford: Johns Hopkins University Press, 2006
Turner A., Antón M. The big cats and their fossil relatives: an illustrated guide to their evolution and natural history. — New York: Columbia University Press, 1997