Проехидны — Zaglossus
Клада Млекопитающеобразные (Mammaliaformes)
Класс Млекопитающие (Mammalia)
Подкласс Йинотерии (Yinotheria)
Клада Австралосфениды (Australosphenida)
Надотряд Прототерии (Prototheria)
Отряд Однопроходные (Monotremata)
Надсемейство Орниторинхоиды (Ornithorhynchoidea)
Семейство Ехидновые (Tachyglossidae)
Род Проехидны (Zaglossus)
Вид Проехидна Брюйна (Zaglossus bruijnii)
Вид Проехидна Бартона (Zaglossus bartoni)
Вид Проехидна Аттенборо (Zaglossus attenboroughi)
? Вид "Проехидна" массивная ("Zaglossus" robusta)
? Вид "Проехидна" Рамсая ("Zaglossus" ramsayi)
Проехидны (Zaglossus Gill, 1877) — род семейства ехидновых (Tachyglossidae Gill, 1872), происходящий из позднего плейстоцена - современности Новой Гвинеи и, вероятно, Австралии. Установлены 3 современных вида. Также род может включать 2 ископаемых вида из плиоцена - плейстоцена Австралии.

Проехидна (Zaglossus sp.).
Синонимы: Tachyglossus Illiger, 1811 (частью); Acanthoglossus Gervais, 1877; Proechidna Gervais, 1877; Bruynia DuBois, 1881; Bruijniia Thomas, 1883; Prozaglossus Kerbert, 1912; ? Megalibgwilia Griffiths et al., 1991.
История изучения
Первое описание проехидны, названной Tachyglossus bruijnii, было сделано в 1876 г. куратором Берлинского зоологического музея профессором В. Петерсом и его коллегой из музея Генуи графом Дориа. Как следует из названия, она была отнесена ими к роду ехидн, а свой видовой эпитет получила в честь голландского купца и натуралиста А.А. Брюйна, благодаря которому остатки этого животного впервые попало в руки ученых. Находясь в горах Арфак на севере Новой Гвинеи, он получил от местного охотника неполный череп (без нижней челюсти), который и был отправлен в Италию.
Уже в 1877 г. Джилл, изучив морфологические особенности проехидны, определил ее в отдельный род Zaglossus (от древнегреческих слов za — "большой" и glossus — "язык"). В последующие годы проехидн неоднократно описывали под все новыми родовыми и видовыми наименованиями, конкурировавшими между собой. С другой стороны, ряд исследователей считал выделение этих таксонов преждевременным. К середине XX в. последняя точка зрения возобладала и сложилась обратная тенденция сводить все известные виды в Zaglossus bruijnii. Лишь в 1998 г. Фланнери и Гровез опубликовали работу, в которой достаточно убедительно обосновали выделение 3 самостоятельных видов и целого ряда подвидов проехидн, отличающихся друг от друга географическим распространением, размерами тела, деталями строения черепа и количеством пальцев на передних и задних конечностях.
Строение и функции
Представители рода более архаичны, чем австралийская ехидна (Tachyglossus aculeatus). Обычно они крупнее последней: длина тела составляет 30-76 см. Вес варьирует в пределах 2-10 кг, а у одной содержавшейся в неволе особи достигал 16,5 кг. Рыльце сильно вытянуто, достигает до 10,5 в длину и занимает 2/3 длины головы. Оно отчетливо загибается книзу, образуя ярко выраженный выпуклый профиль, тогда как рыльце австралийской ехидны короче и прямее. В степени изогнутости рыльца наблюдается значительная внутривидовая изменчивость. На его конце расположены ноздри и маленький рот. По бокам задней части головы имеются небольшие ушные раковины.

Вид Проехидна Бартона (Zaglossus bartoni).
По мере взросления рыльце проехидны удлиняется и становится более массивным в своей передней части; носовое отверстие с возрастом укорачивается и сзади закругляется; кости заднего неба увеличивают свою массивность, а их края становятся заметными ниже основания черепа при взгляде сзади; большое затылочное отверстие, обычно широко открытое к началу половозрелости, в дальнейшем постепенно сужается; кости черепа в большинстве случаев сливаются друг с другом, в числе последних исчезают швы между носовыми и лобными костями. Костное небо является самым длинным среди известных млекопитающих.
В позвоночнике проехидн 43 позвонка — 7 шейных, 17 грудных, 4 поясничных, 3 крестцовых и 12 хвостовых (возможно, их число в грудном и крестцовом отделах варьирует). Туловище широкое, с куполообразной спиной и плоским брюхом. Передние и задние конечности примерно вдвое выше, чем у ехидны, с хорошо развитой мускулатурой. Возможно, более длинные ноги связаны с необходимостью лучшего обзора в густом подлеске. I-й и V-й пальцы кисти и стопы могут редуцироваться вплоть до небольших мозолистых бугорков, лишенных когтей. На кисти крайние пальцы редуцируются в большей степени, чем на стопе. Пальцы с II-го по IV-й всегда вооружены мощными когтями, которые на задних конечностях постепенно уменьшаются в направлении изнутри кнаружи. У взрослых самцов на внутренней поверхности пятки задней лапы имеется хорошо развитая роговая шпора, которые у самок исчезает еще до наступления половой зрелости. Шпора самца связана каналом со слабо развитой подколенной железой. Хвост развит лучше чем у ехидны, его длина достигает 5-7 см.
Иглы короче (3-5 см длиной), жестче, имеют меньшие внутренние полости и расположены реже, чем у австралийской ехидны. Иногда редкие иголки есть даже на брюшной поверхности. Окрас игл варьирует от почти белого и светло-серого до насыщенного черного, с темными или светлыми кончиками. Наблюдается значительное разнообразие в длине и характере размещения их на теле. Жесткие, густые и длинные волосы часто полностью скрывают иглы на спине. Окраска шерсти варьирует от светло-коричневой до черной; на голове она обычно более светлая, чем на туловище. Некоторые особи имеют бледные, почти белые отметины на морде, конечностях и крестце. Зафиксированы альбиносы. В период выведения потомства у самок на задней части брюха разросшиеся складки кожи образуют слабо выраженную сумку.
Строение языка существенно отличается от такового у австралийской ехидны. Он способен выдвигаться из ротового отверстия на 30 см, но обычно высовывается лишь на 2-3 см. В языке отсутствует кольцевая мускулатура. Сверху в передней 1/3 языка залегает глубокая бороздка, в передних 3/4 которой расположены 3 продольных ряда очень острых и достаточно крупных, направленных назад роговых шипов. 1 ряд шипов находится на дне бороздки и 2 ряда — по бокам. Шипы в них чередуются: сперва идет шип нижнего ряда, затем боковых и т. д. При высовывании язык загибается вниз, раскрывая бороздку. Во втянутом состоянии бороздка замкнута. Также на передней части языка есть более мелкие изогнутые шипы, расположенные на ней снизу, по бокам и по сторонам вверху, но отсутствующие на середине верхней поверхности языка и внутри бороздки. Эти шипы гомологичны шипам на нижней стороне языка ехидны. Наконец, сзади на верхней части языка расположена подушечка из мелких, ороговевших, направленных назад шипов. Пища измельчается благодаря тому, что они трутся о противостоящие им шипы на небных складках. Данная конструкция соответствует таковой у ехидны, но развита в меньшей степени и с экзоскелетами членистоногих справляется плохо. Общее количество шипов в ротовой полости больше, чем у ехидны. Слюнные железы хорошо развиты.
Как и у ехидны, чрезвычайно тонкое обоняние играет первостепенную роль при выслеживании обитающей в почве добычи. Этой же цели служат более 2000 электрорецепторов, расположенных со средней плотностью 12 на 1 кв. мм в толще кожи на дистальной части рыльца и снабженных нервными окончаниями. Кроме того, имеется большое количество механорецепторов. Неплохо развит у проехидн и слух. А вот их зрение не отличается остротой.
У проехидн в большей степени, чем у других однопроходных, проявляется гетеротермность. Температура их тела, в зависимости от температуры окружающей среды, может меняться от 34,2 до 24,2 °C, составляя в среднем около 30 °C. При этом животные продолжают оставаться активными и погружаются в кратковременное оцепенение лишь при самых неблагоприятных условиях. В толще коже залегают апокриновые потовые железы. Средняя интенсивность обмена веществ составляет 6,5 Вт. Основной уровень метаболизма равен 25,5 кДж/час. Потребление кислорода особью весом 10,3 кг находилось на уровне 0,118 мл/г/час.
В кариотипе 63 (самец) или 64 (самка) хромосомы.
Распространение и среда обитания
Относительно широко распространены на Новой Гвинее, но на большей части ареала плотность популяции невысока. Тяготеют к отдаленным и труднодоступным районам, редко посещаемым человеком. Встречаются от низменностей до горных вершин высотой 4150 м над ур. моря. Населяют прежде всего влажные тропические леса (примерно до 3000 м над ур. моря), субальпийские и альпийские луга (приблизительно от 3000 м над ур. моря), а также кустарниковые заросли и каменистые участки. Местностей с недостатком влаги, равно как и болот, избегают.
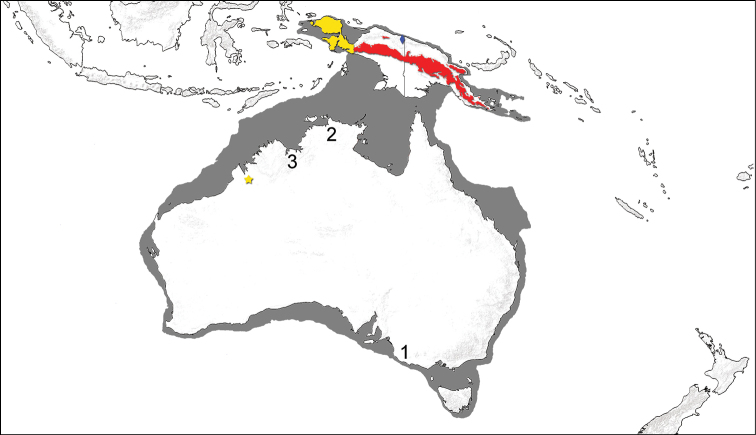
Карта суперматерика Сахул, существовавшего до конца последнего оледенения и через обнаженный континентальный шельф (показан темно-серым) объединявшего Австралию с Новой Гвинеей, Тасманией и другими прилегающими островами. Желтым цветом отмечен современный ареал проехидны Брюйна (Zaglossus bruijnii), красным — проехидны Бартона (Zaglossus bartoni), синим — проехидны Аттенборо (Zaglossus attenboroughi). Желтой звездочкой отмечено место поимки проехидны Брюйна в округе Кимберли, Западная Австралия. Другие возможные следы присутствия проехидн на территории Австралии: 1 — ископаемые остатки плейстоценового возраста из группы пещер Наракурт, Южная Австралия; 2 — голоценовый или позднеплейстоценовый наскальный рисунок на полуострове Арнем-Ленд, Северная Территория; 3 — сообщения аборигенов из Кунунарры, Западная Австралия, о встречах с проехидной в XX в.
Состав
К роду относят 3 ныне живущих вида.
Проехидна Брюйна (Zaglossus bruijnii Peters et Doria, 1876) — типовой вид, известный из современности запада Новой Гвинеи и, по-видимому, Западной Австралии.
Проехидна Бартона (Zaglossus bartoni (Thomas, 1907)) — вид, известный из современности центра и востока Новой Гвинеи.
Проехидна Аттенборо (Zaglossus attenboroughi Flannery et Groves, 1998) — вид, известный из современности севера Новой Гвинеи.

Формы проехидн (Zaglossus).
Остатки ископаемых форм обычно настолько фрагментарны, что их видовая принадлежность во многом остается сомнительной. Более того, само их включение в род проехидн пока является лишь предварительным. Дело также осложняется неопределенностью датировок. Вероятно, вымирание этих крупных однопроходных на материковой части Австралии и в Тасмании было вызвано позднеплейстоценовым иссушением климата, которое привело к значительному сокращению площади лесов и влажных лугов, в мягкой почве которых в изобилии встречались поедаемые земляные черви и личинки насекомых. Пригодная для проехидн среда обитания сохранилась главным образом на Новой Гвинее. В периоды колебаний температуры и влажности вертикальная зональность, характерная для склонов здешних гор, обеспечивала подходящие жизненные условия независимо от климатических сдвигов. На Новой Гвинее проехидны обитают и сейчас. В других рефугиумах — на севере Австралии и в Тасмании — они уже исчезли.
? "Проехидна" массивная ("Zaglossus" robusta Murray, 1978) (синонимы: Ornithorhynchus maximus Dun, 1894; Echidna (Proechidna) robusta Dun, 1894; Megalibgwilia robusta Griffiths et al., 1991) — вид, известный из плейстоцена или плиоцена Нового Южного Уэльса. Нередко определяется в род мегалибвилий (Megalibgwilia).
? "Проехидна" Рамсая ("Zaglossus" ramsayi Griffiths, Wells et Barrie, 1991) (синонимы: Echidna oweni Kreft, 1868; Echidna ramsayi Owen, 1884; Zaglossus harrisoni Scott et Lord, 1922; Megalibgwilia ramsayi Griffiths et al., 1991) — вид, известный из плиоцена - голоцена Западной Австралии, Квинсленда, Южной Австралии, Нового Южного Уэльса, Виктории, острова Кинг в Бассовом проливе, а также Тасмании. Зачастую относится к роду мегалибвилий (Megalibgwilia).
Zaglossussp. Piper, 2007 — форма, установленная из раннего плейстоцена Нового Южного Уэльса, отложения формации Нельсон-Бэй.
Zaglossus sp. — форма, установленная из позднего плейстоцена Южной Австралии.
Zaglossus sp. — форма, установленная из позднего плейстоцена Новой Гвинеи.

Наскальный рисунок с полуострова Арнем-Ленд (Северная Территория, Австралия), предположительно изображающий проехидну. Он выполнен в древнем одноцветно-многослойном стиле и датируется поздним плейстоценом или ранним голоценом. Об этом свидетельствуют обнаруженные в той же местности наскальные рисунки сумчатого дьявола (Sarcophilus laniarius) и сумчатого волка (Thylacinus cynocephalus), вымерших на континенте несколько тысяч лет назад.
Географическое распространение проехидн представляет для таксономистов особенно сложную проблему. Вопрос о числе выделяемых форм также усложняется ввиду значительной возрастной, половой и индивидуальной изменчивости. В настоящее время принято считать, что идентификационные различия в строении черепа и числе когтей обеспечивают достаточное доказательство того, что проехидны, живущие в изолированных друг от друга популяциях к западу и востоку от озера Паниаи, представляют собой разные виды. Вполне возможно, что у них был общий регион происхождения, но подтверждающие это ископаемые данные пока отсутствуют.
Образ жизни
Проехидны весьма скрытные животные, поэтому их повадки изучены плохо. Активны они главным образом в сумерках или ночью, а днем спят. Передвигаются довольно медленно, со скоростью около 1 км/ч. Бредут шаркающей походкой, с низко опущенной головой, уткнувшись рыльцем в землю. Если на пути попадается большой камень или бревно, животное скорее перелезет через него, чем обойдет, а лужу или озерцо — переплывет. В неволе проехидны отлично перелезают через перегородки высотой до 30 см и подтягиваются передними лапами на высоту 45 см и более. Воды они тоже не боятся, охотно используют ее для чистки, бродят по мелководью и даже полностью погружаются в водоем, исследуя дно рыльцем и собирая с него пищу. При этом проехидны надолго задерживают дыхание. Могут подолгу отдыхать, лежа в емкости с водой и периодически выставляя рыльце для того, чтобы подышать.
Также длинное рыльце активно используется в качестве дополнительной конечности, позволяющей преодолевать различные препятствия. Например, в неволе животное влезало на стул. Вначале оно вставало на задние ноги, упираясь передними в ножки стула. Затем, цепляясь когтями передних лап и рыльцем за сидение, постепенно скользило вверх задними ногами по ножкам стула, захватывая при этом все больший участок сидения. Слезая со стула, проехидна сначала пробовала высоту задними ногами, затем опускала голову и пыталась достать рыльцем до пола. Потом животное пыталось измерить расстояние до окружающих предметов и до пола языком, высовывая его на полную длину. В конце концов проехидна плавно сползала со стула головой вниз, удерживая себя задними ногами, и приземлялась на голову, подгибая рыльце к брюху. Помимо этого, рыльце может выступать в качестве рычага — с его помощью животные переворачивают и сдвигают камни диаметром до 25 см. В поисках корма проехидны прощупывают рыльцем лесную подстилку и рыхлый верхний слой грунта, полагаясь в основном на хорошо развитый нюх. В результате в почве образуются хорошо узнаваемые углубления в форме рыльца и передней части головы, которые могут достигать 40 см в глубину. После обнаружения добыча быстро выкапывается когтистыми передними лапами.
Рацион состоит в основном из земляных червей, многочисленных во влажной почве Новой Гвинее и достигающих здесь крупных размеров. Обнаружив скрытого в лесной подстилке достаточно крупного червяка, проехидна сначала тыкается в него рыльцем, пытаясь найти головной или хвостовой конец. Если червь сильно извивается, она наступает на него лапой, и продолжает искать конец. Располагая рыльце вертикально над концом червя, животное высовывает конец изогнутого вниз языка с раскрытой бороздкой и подцепляет червя находящимися в ней шипами, слегка двигая рыльцем вперед и вверх. Затем червь втягивается в рот при помощи ретракции языка, сопровождающейся серией подергиваний. По-видимому, эти подергивания отображают перехватывание червя более дальними шипами, пока он не достигнет перетирающего аппарата. Звук перетирания слышим, как у ехидны. Дополнительно поедаются мелкие многоножки, личинки жуков, бабочек и цикад, а также небольшое количество муравьев и термитов (впрочем, при содержании в неволе некоторые особи муравьев вообще не употребляют). Такая добыча просто слизывается с субстрата кончиком языка. В поисках пищи проехидны не только роются в земле, но и вскрывают когтями гнилые бревна. В неволе охотно едят мелко измельченные или порезанные на тонкие полоски сырое мясо и печень, находя по запаху предложенную пищу на расстоянии не менее 1,5 м. Могут голодать в течение нескольких недель. Пьют, погрузив рыльце в воду и втягивая ее в рот с шумом и пузырями.
Если проехидну напугать, она прячется в укрытие или приседает, подгибая под себя рыльце и выставив наружу иглы. Убежищами им служат норы, кустарниковые заросли, завалы валежника и нагромождения камней. Нора обычно устраивается в склоне (так животному легче копать) среди маскирующей густой растительности, а ее длина может достигать нескольких метров.Вне периода размножения ведут одиночный образ жизни. Известно, что каждая взрослая особь занимает определенную территорию. Проехидны способны издавать сопение и негромкие фыркающие звуки. В остальном их социальная биология практически не изучена.
Сегодня эндемичные млекопитающие хищники, охотящиеся на проехидн, отсутствуют. В прошлом таковым мог являться сумчатый волк (Thylacinus cynocephalus), исчезнувший на Новой Гвинее несколько тысяч лет назад. Иногда на этих животных нападает одичавшая домашняя собака (Canis lupus familiaris), завезенная на остров древними мореплавателями.
Жизненный цикл
Данные о размножении проехидн крайне ограничены. Считается, что в этом отношении они сходны с лучше изученной австралийской ехидной. Размножаются 1 раз в году. Брачный сезон ориентировочно приходится на июль. О взаимодействиях самцов и самок в период спаривания ничего неизвестно. Длительность беременности предположительно составляет 3-4 недели. Самка откладывает 1 небольшое яйцо (по неподтвержденным сведениям, полученных от местных охотников — до 3-6 яиц), помещаемое ею в сумку. Спустя приблизительно 1,5-2 недели из яйца вылупляется детеныш. Примерно до 6 месяцев он питается молоком, которое выделяется на железистых полях внутри сумки по обе ее стороны. Насколько известно, яиц и переносимых в сумке детенышей никто из специалистов пока не видел и в музейных коллекциях они отсутствуют.
Продолжительность жизни в природе не установлена, в неволе одна особь прожила по меньшей мере 48 лет и 5 месяцев. Это примерно вдвое больше, чем можно было бы ожидать исходя из размеров тела этих животных. Длительность одного поколения оценивается в 16 лет.

Проехидна Брюйна (Zaglossus bruijnii), содержащаяся в неволе.
Угрозы и охрана
Из всех ныне живущих однопроходных проехидны находятся в самом угрожаемом положении и крайне нуждаются в защите. Большой ущерб их популяции наносит вырубка первичных лесов ради ценной древесины и под сельскохозяйственные угодья. В результате среда обитания становится гораздо более доступной для охотников, которые добывают проехидн ради их жирного мяса. Последнее напоминает по вкусу свинину и, обжаренное в углях костра, высоко ценится местными жителями. В прошлом истребление проехидн частично сдерживали табу, но после вторжение западной системы ценностей традиционные верования практически повсеместно отмерли. В некоторых районах в охотничьих целях даже применяются специально обученные собаки, находящие животных по запаху в их хорошо скрытых убежищах. Несмотря на то, что спортивная охота на проехидн в Западной Новой Гвинее (Индонезия) и на Папуа-Новой Гвинее официально запрещена, традиционная охота на них по-прежнему ведется. От преследования особенно страдают проехидны, живущие в низменностях, где плотность человеческой популяции выше. С другой стороны, населенные этими животными леса на возвышенностях во время длительных засух страдают от пожаров.
Согласно статистической оценке, в 1982 г. насчитывалось порядка 300 тыс. проехидн, плотность популяции которых составляла 1,6 особи на 1 кв. км ареала. Эти цифры, полученные экстраполяцией, явно завышены. Сегодня все представители рода признаны находящимися под угрозой исчезновения. Поскольку о распространении, биологии и экологии этих животных мало что известно, затруднительно выработать эффективный комплекс мер по их защите и сохранению. Скрытность и ночной образ жизни проехидн сильно затрудняют их поиск и изучение, а иглистый покров осложняет прикрепление к телу следящих устройств.
Проехидны содержатся в ряде зоопарков, где они живут подолгу, хотя и не размножаются. Наблюдения, сделанные в неволе, позволили получить ряд интересных и ценных сведений. Для того, чтобы посетители могли наблюдать за активностью проехидн в дневное время, в некоторых зоопарках искусственно смещается суточный цикл.
Ссылки
Long-beaked echidna (Wikipedia) перейти
Zaglossus (Paleobiology Database) перейти
Литература
Жизнь животных. В 7 т. Т. 7: Млекопитающие / Гл. ред. Соколов В. Е. — М.: Просвещение, 1989
Жизнь животных по А. Э. Брему. В 5 т. Т. 5: Млекопитающие / Общ. ред. Северцов А. Н. — М.: Государственное учебно-педагогическое издательство Наркомпроса РСФСР, 1941
Кист А. Австралия и острова Тихого океана. — М.: Прогресс, 1980
Кістяківський О. Б., Корнєєв О. П. Посібник з зоогеографії. — К.: Радянська школа, 1968
Кэрролл Р. Палеонтология и эволюция позвоночных: В 3-х т. Т. 3. — М.: Мир, 1993
Млекопитающие: Полная иллюстрированная энциклопедия. В 2 кн. / Ред. Макдональд Д. В. — М.: Омега, 2007
Павлинов И. Я. Систематика современных млекопитающих. — М.: МГУ, 2006
Россолимо О. Л., Павлинов И. Я., Крускоп С. В., Лисовский А. А., Спасская Н. Н., Борисенко А. В., Панютина А. А. Разнообразие млекопитающих. Ч. 1-3. — М.: Изд-во КМК, 2004
Соколов В. Е. Редкие и исчезающие животные. Млекопитающие. — М.: Высшая школа, 1986
Соколов В. Е. Систематика млекопитающих. Отряды: однопроходных, сумчатых, насекомоядных, шерстокрылов, рукокрылых, приматов, неполнозубых, ящеров. — М.: Высшая школа, 1973
Dawson L. Marsupial fossils from Wellington Caves, New South Wales; the Historic and Scientific Significance of the Collections in the Australian Museum, Sydney. Records of the Australian Museum 37 (2), 1985
Flannery T. F. Mammals of New Guinea. — Ithaca: Cornell University Press, 1995
Flannery T. F., Groves C. P. A revision of the genus Zaglossus (Monotremata, Tachyglossidae), with description of new species and subspecies. Mammalia 62 (3), 1998
Griffiths M. The Biology of the Monotremes. — New York - London: Academic Press, 1978
Grzimek's animal life encyclopedia. Volumes 12-16 / Ed. by Hutchins M., Kleiman D. G., Geist V., McDade M. C. — 2rd ed. — Farmington Hills, MI: Gale Group, 2004
Helgen K. M., Miguez R. P., Kohen J., Helgen L. Twentieth century occurrence of the Long-Beaked Echidna Zaglossus bruijnii in the Kimberley region of Australia. ZooKeys 255, 2012
Mammal Species of the World. A Taxonomic and Geographic Reference / Ed. by Wilson D. E., Reeder D. M. — 3rd ed. — Baltimore: Johns Hopkins University Press, 2005
Murray P. F. Furry egg-layers: the monotreme radiation. In Archer M., Clayton G. (eds). Vertebrate Zoogeography and Evolution in Australasia. — Perth: Hesperian Press, 1984
Musser A. M. Diversity and relationships of living and extinct monotremes. Australian Mammalogy 21, 1999
Musser A. M. Furry Egg-layers: Monotreme Relationships and Radiations. In Merrick J. R., Archer M., Hickey G. M., Lee M. S. Y. (eds). Evolution and Biogeography of Australasian Vertebrates. — Sydney: Australian Scientific Publishing, 2006
Nowak R. M. Walker’s Mammals of the World. Monotremes, Marsupials, Afrotherians, Xenarthrans, and Sundatherians. — Baltimore: Johns Hopkins University Press, 2018
Nowak R. M., Paradiso J. L. Walker’s Mammals of the World. — 4nd ed. — Baltimore/London: Johns Hopkins University Press, 1983
Rismiller P. D., Grutzner F. Tachyglossus aculeatus (Monotremata: Tachyglossidae). Mammalian Species 51 (980), 2019
Van Deusen H. M. Zaglossus: egg-laying anteater. Fauna 2, 1971