

Репеномамы представляли собой крепко сложенных млекопитающих с большой головой и вытянутым туловищем, покоящимся на относительно коротких конечностях.
Череп массивный, с тяжелой нижней челюстью. Зубная формула: I3/i2, Cl/c1, Р2/p2, М4/m5. Верхние и нижние резцы крупные. Верхний клык расположен у шва между предчелюстной и верхнечелюстной костями и по форме сходен с резцами. Предкоренные зубы конические с заостренной вершинкой. Коренные зубы уменьшаются в размере кзади, с 3 закругленными вершинками. m5 прорезывался последним, уже во взрослом возрасте. Как это характерно для млекопитающих, постоянные щечные зубы немного превышают по размерам своих молочных предшественников. Зубы взрослых особей имеют выраженные следы износа. В целом, зубочелюстной аппарат выраженно плотоядного типа.
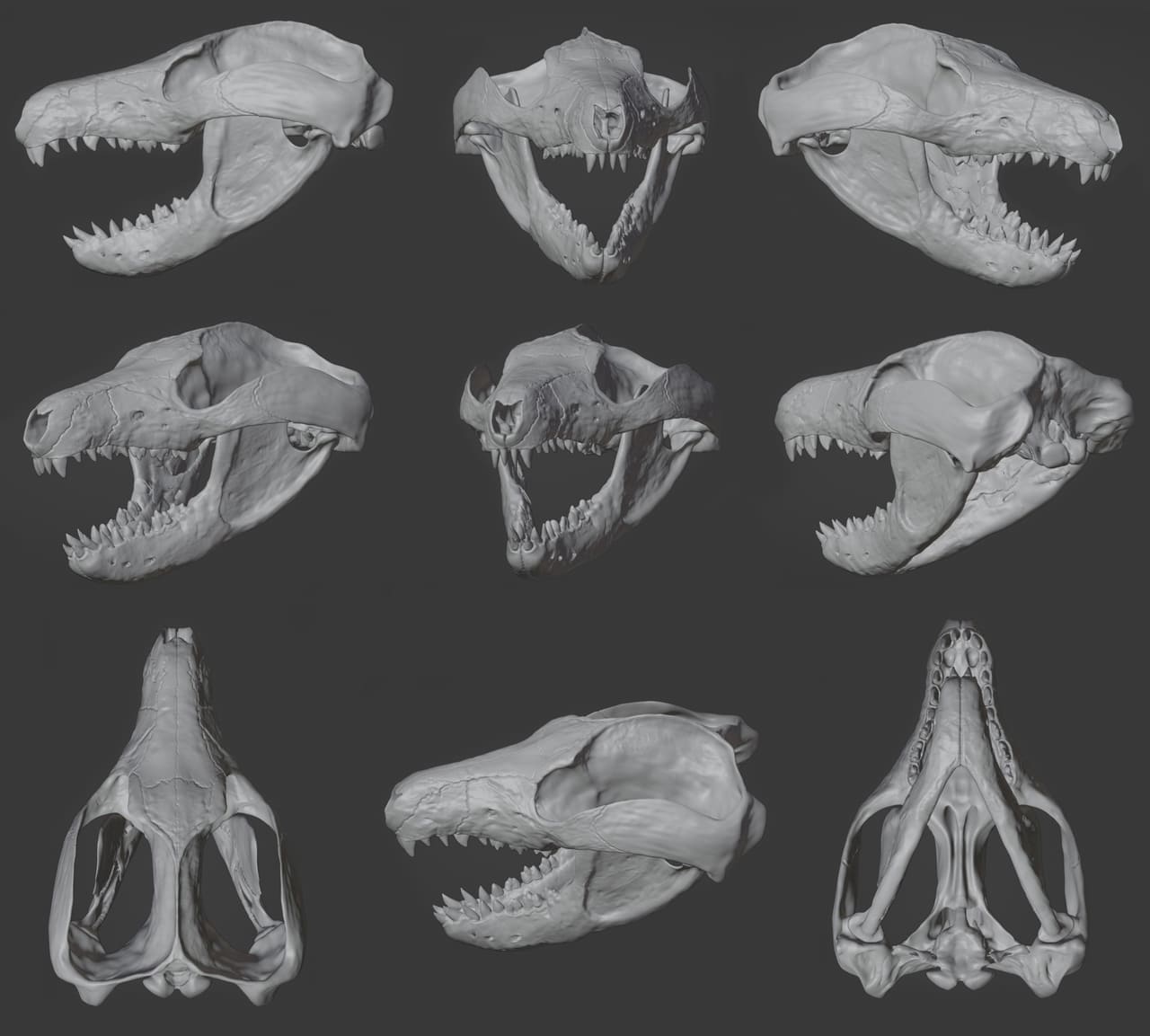
Грудной и поясничный отделы позвоночника хорошо дифференцированы. Лопатка имеет большую ость и суставную ямку, обращенную вентрально. Головка плечевой кости полусферическая, отклоняется заднедорсально и закручена под углом около 25 ° по отношению к дистальному концу. Головка бедренной кости отклоняется от диафиза дорсомедиально и обращена кпереди. Медиальный мыщелок дистального отдела бедренной кости уже и глубже латерального, оба направлены задневентрально. Плечевая и бедренная кости отходят от суставов плечевого и тазового поясов под широким углом. Крупный локтевой отросток и задневентрально направленные мыщелки бедренной кости позволяли конечностям занимать полувыпрямленное положение, что характерно и для других примитивных териеобразных млекопитающих. Кисть и стопа короткие и широкие, стопоходящего типа. Судя по всему, репеномамы не могли быстро бегать.


Массивный репеномам (Repenomamus robustus Li, Wang, Wang et Li, 2000) (тип) — типовой вид, известный из раннего мела (пачка отложений Луцзятунь, базальная часть формации Исянь, низы аптского яруса) окрестностей деревни Луцзятунь в провинции Ляонин, Китай. Череп достигал около 7 см в длину, общая длина тела составляла порядка 65 см, а вес оценивается в 4-6 кг, т. е. животное было размером с виргинского опоссума (Didelphis virginiana).
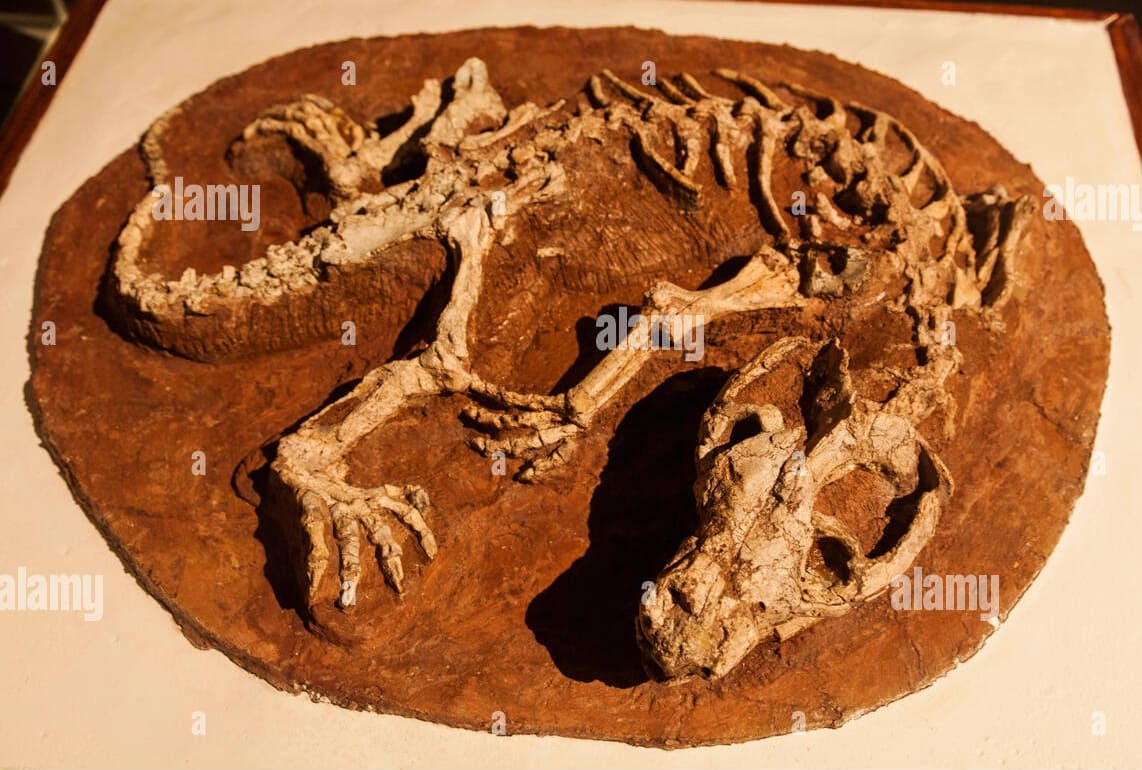


Сохранившиеся зубы, кости конечностей и фаланги подтверждают, что эти кости принадлежат ювенильному пситтакозавру. Длина его головы и туловища оценивается в 14 см, что составляет около 1/3 длины головы и туловища Repenomamus robustus. Большинство зубов изношены — это свидетельствует о том, что скелет пситтакозавра не является эмбрионом, добытым из яйца. Некоторые длинные кости конечностей сохранились в сочленении, но череп и другие кости раздроблены на фрагменты, что позволяет предположить, что ювенильный Psittacosaurus lujiatunensis был разорван и проглочен кусками. Судя по многочисленным скоплениям остатков пситтакозавров, их молодые особи обычно держались группами рядом с родителями, поэтому представляется вероятным, что репеномам вылавливал молодняк, когда он разбредался, или, возможно, совершал набеги на их гнезда.
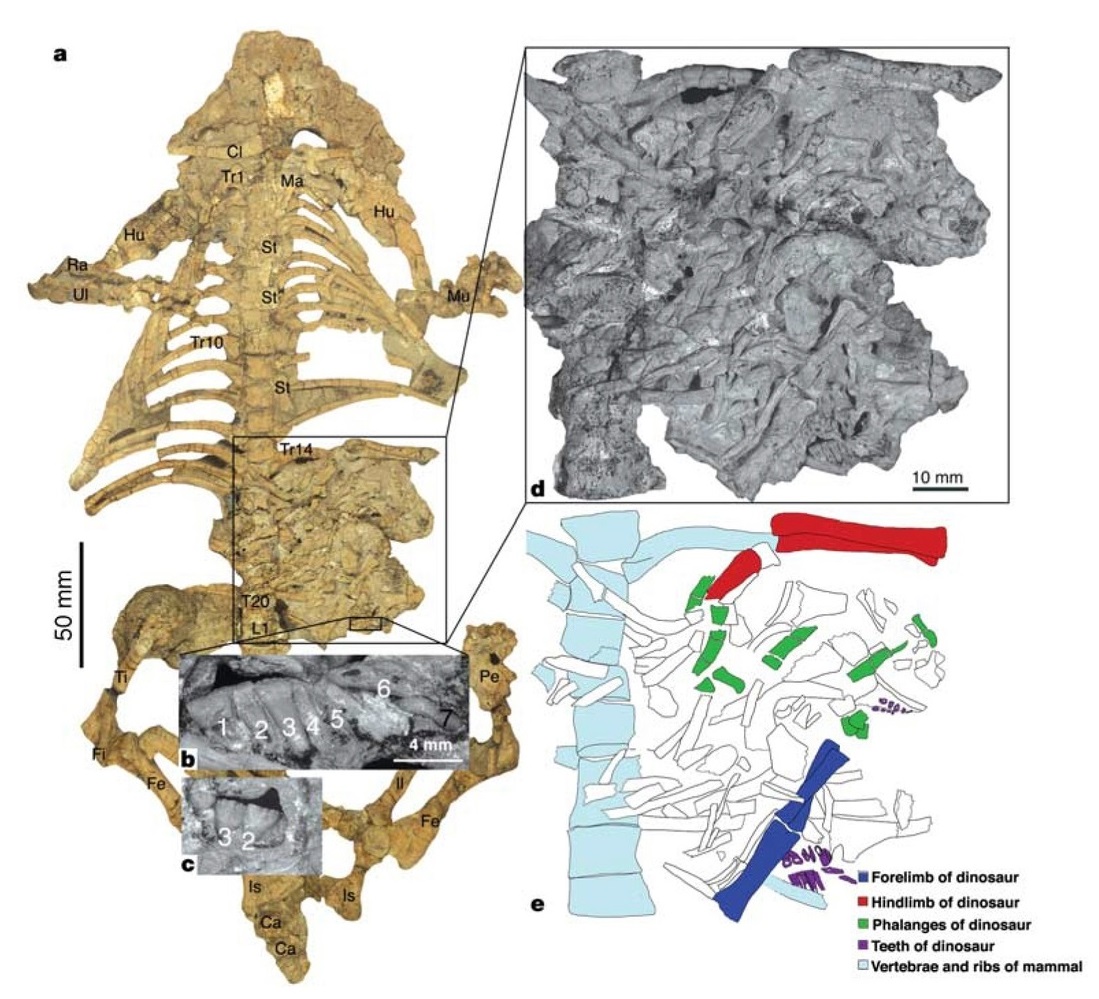

Почти идеальная сохранность скелетов обоих животных исключают возможность того, что их остатки смешались уже после гибели в результате тафономических процессов. Эти процессы действительно привели к смещению некоторых элементов, в числе которых нижняя челюсть и прокушенные передние ребра пситтакозавра, а также конечные фаланги пальцев и конечные хвостовые позвонки репеномама, но в остальном скелеты дошли до нас почти нетронутыми. Тела животных явно не переносились на значительное расстояние и были захоронены на месте гибели, причем оба события происходили если не одновременно, то близко по времени.
Авторы открытия полагают, что Repenomamus robustus активно атаковал более крупного Psittacosaurus lujiatunensis, когда оба животных попали в вулканический грязевой поток и были им завалены. Ученые признают, что репеномам мог поедать тушу уже погибшего пситтакозавра, однако они отмечают, что на костях динозавра отсутствуют следы зубов других хищников или падальщиков. Кроме того, если бы репеномам поедал уже мертвого ящера, он вряд ли стал бы забираться на него сверху, и тела животных вряд ли бы переплелись таким образом, как это наблюдается на окаменелости. Таким образом, более вероятно, что репеномам повалил пситтакозавра (по всей видимости, впавшего в шоковое состояние и переставшего бороться за жизнь) и начал его поедать, после чего хищник и жертва были внезапно погребены под слоем жидкой грязи.
Питание живой добычей часто наблюдается у ныне живущих плотоядных млекопитающих, включая гиеновых собак (Lycaon pictus), пятнистых гиен (Crocuta crocuta) и шакалов (Canis mesomelas, Canis aureus). Фактически, после первоначальной борьбы их жертва может отказаться от самозащиты, предпочтя вместо этого пассивно лечь, находясь в состоянии истощения и глубокого шока. Кроме того, клептопаразитизм крупных хищников в африканской саванне может существенно изменить охотничьи и кормовые привычки более мелких плотоядных, и поедание еще живой добычи гиеновыми собаками, возможно, является одной из таких адаптаций. Более крупные тероподы раннемеловой экосистемы Луцзятунь могли представлять равную угрозу для репеномамов, мотивируя их аналогичное хищническое кормовое поведение.
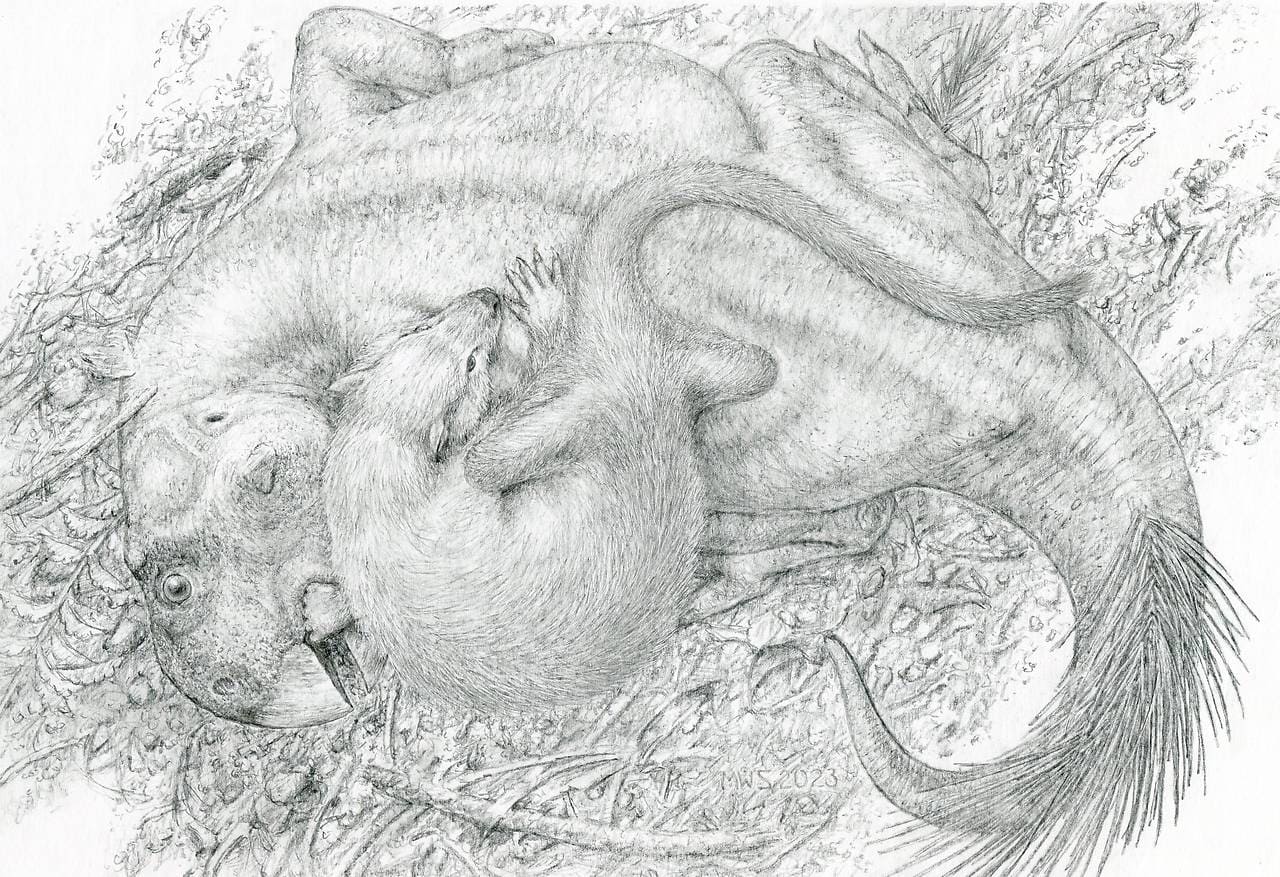
Хотя может показаться, что пситтакозавр — слишком крупная добыча для репеномама, среди современных хищников тоже есть те, что охотятся на жертв, значительно превышающих их по размеру. Например, ласка (Mustela nivalis) время от времени нападает на гораздо более крупных глухарей (Tetrao spp.), рябчиков (Tetrastes bonasia) и зайцев (Lepus spp.), а росомаха (Gulo gulo) может добывать лосей (Alces alces) и северных оленей (Rangifer tarandus). Более того, авторы исследования предполагают, что даже Psittacosaurus lujiatunensis, достигшие максимального веса в 23,5 кг, оставались уязвимыми для взрослых Repenomamus robustus, которые могли весить до 5,54 кг.
Необходимо отметить, что подлинность данной находки вызывает сомнения у ряда специалистов. Впрочем, они не получили непосредственного доступа к окаменелостям и судят о них лишь по фотографиям. В первую очередь, скептиков смущает идеальная даже по меркам Исяньской формации сохранность упомянутых остатков. Они указывают, что как бы хорошо ни были сохранны скелеты, всегда наблюдаются их деформации и тафономические нарушения. Кроме того, на их взгляд, кости выглядят так, будто они напрямую соприкасаются друг с другом и были лишены плоти в момент контакта. Кроме того, по мнению скептиков, авторы открытия приложили недостаточно усилий для доказательства его подлинности — например, не предоставили рентгенограмму окаменелости, что подтвердило бы целостность составляющих ее элементов.



Гигантский репеномам (Repenomamus giganticus Hu, Meng, Wang et Li, 2005) — вид, известный из раннего мела (пачка отложений Луцзятунь, базальная часть формации Исянь, низы аптского яруса) окрестностей деревни Луцзятунь в провинции Ляонин, Китай. Типовой образец IVPP V14155 представляет собой скелет, включающий большую частью черепа в ассоциации с правой ветвью нижней челюсти с полными правыми зубными рядами и сохранившийся в сочленении посткраниум без кистей, стоп и конца хвоста. Череп составляет 16 см в длину, туловище — 52,2 см, сохранившаяся часть хвоста — 36,4 см. Таким образом, общая длина несколько превышала 1 м, а вес оценивается в 12-14 кг, что сравнимо с крупным сумчатым дьяволом (Sarcophilus laniarius). Животное было на 60 % длиннее и по меньшей мере вдвое тяжелее, чем Repenomamus robustus.

Помимо более значительных общих размеров, этот вид отличается от Repenomamus robustus тем, что его череп имеет более мощные сагиттальный гребень, лямбдовидный гребень и скуловые дуги; резцы пропорционально большие; верхний клык двукорневой; I-й верхний предкоренной намного уступает верхнему клыку по величине; верхние коренные зубы с полным язычным и частичным щечным пояском; нижняя челюсть более массивная, с наклонно ориентированным пропорционально более глубоким симфизом, широким венечным отростком и глубокой жевательной ямкой; резцы, клыки и предкоренные расставлены менее широко; вершинки на нижних коренных развиты лучше.
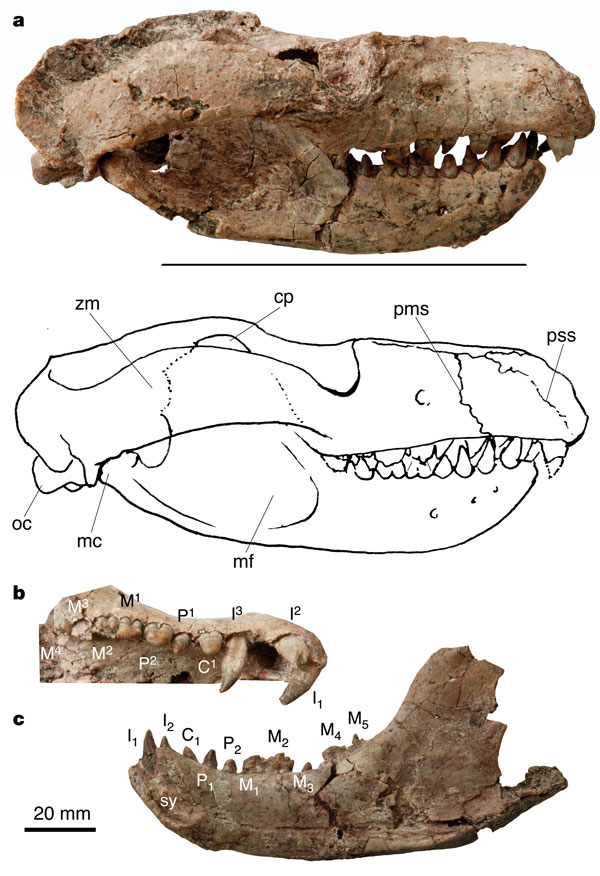

Крупные заостренные резцы и сходные по форме клыки и предкоренные образуют у репеномамов аппарат для захвата, удержания и разрывания добычи. Этот аппарат приводился в действие сильной челюстной мускулатурой, о чем свидетельствуют мощные зубная и скуловая кости, большая ямка височной мышцы и глубокая ямка жевательной мышцы. В то же время, небольшие и тупокоронковые кореннные зубы играли лишь вспомагательную роль в обработке пищи. Наличие крупных кусков добычи в желудке Repenomamus robustus подтверждают, что жевание, вероятно, не было присуще репеномамам. Большие заостренные передние зубы, сопровождаемые мелкими щечными зубами, характерны для многих плотоядных синапсид, не относящихся к млекопитающим.

Находки остатков репеномамов существенно расширили представления об экологических нишах, занимаемых млекопитающими во время господства рептилий. До их открытия мезозойские млекопитающие были известны главным образом как мелкие зверьки, предположительно ведущие ночной образ жизни. Причины их небольших размеров точно не установлены, но обычно считалось, что ниши животных размером около 1 м и более заполняли исключительно динозавры, крокодилы, черепахи и другие рептилии (преимущественно, дневные) вплоть до их вымирания на рубеже мелового и палеогенового периодов и широкой диверсификации млекопитающих в кайнозое. Репеномамы максимально приблизились к верхнему предел размеров тела мезозойских млекопитающих и фактически превосходят по величине некоторых живших по соседству хищных динозавров, с которыми они могли успешно конкурировать за кормовые ресурсы. Более крупные животные способны жить дольше и передвигаться быстрее, но им также требуется большее количество пищи. Судя по размерам тела, Repenomamus giganticus мог охотиться на более крупную добычу на более обширном участке, чем Repenomamus robustus.
