Сфенакодонты — Sphenacodontia
Класс †Синапсиды (Synapsida)
Подкласс †Пеликозавры (Pelycosauria)
Надотряд †Сфенакообразные (Sphenacomorpha)
Отряд †Сфенакодонты (Sphenacodontia)
Сфенакодонты (Sphenacodontia Williston, 1912) — группа синапсид, рассматриваемая в составе подотряда эвпеликозавров (Eupelycosauria Kemp, 1982) согласно классической систематике Р. Рейса, либо в составе клады сфенакообразных (Sphenacomorpha Ivakhnenko, 2003) в альтернативной систематике М.Ф. Ивахненко. Происходят из позднего карбона - ранней перми Европы и ранней - ? средней перми Северной Америки. Установлены безранговая клада и 2-3 семейства, объединяющие в общей сложности порядка 20 родов.
Реконструкции внешнего облика секодонтозавра (Secodontosaurus obtusidens) (вверху) и диметродона (Dimetrodon grandis) (внизу). Автор — Г. Угуэто.
Размеры небольшие, средние и крупные: общая длина тела варьирует от 0,6 до 4,5 м.
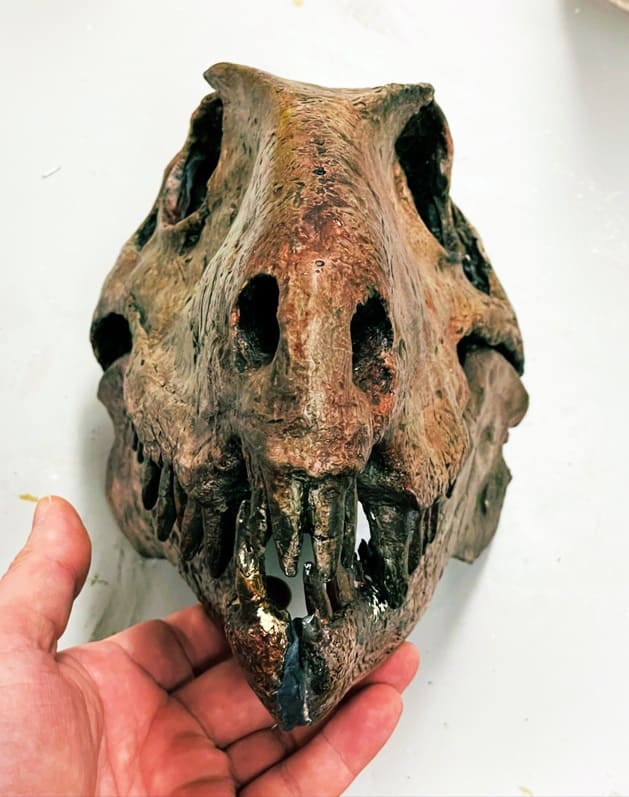
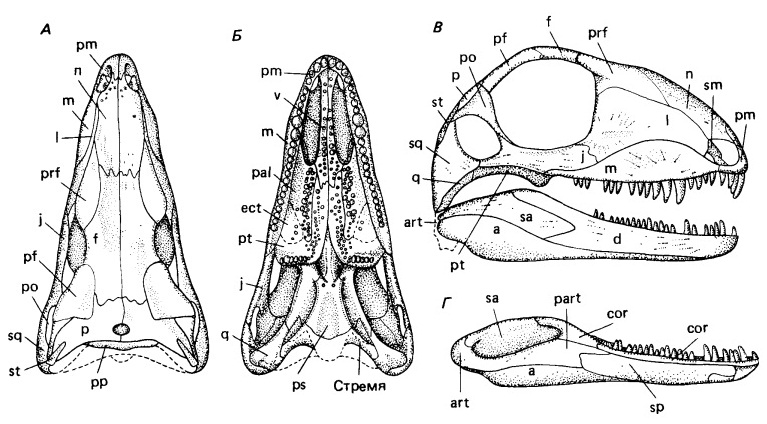
Череп крупный относительно тела, с умеренно удлиненной мордой и высокий (у Haptodontinae и Sphenacodontinae), либо низкий с крайне удлиненной и узкой мордой (у Secodontosaurinae). На уровне глазниц череп суживается и снова расширяется в своей задней части. Слезная кость достигает ноздри (у Haptodontinae), либо укорачивается и не достигает (у Secodontosaurinae и Sphenacodontinae). У многих сфенакодонтов, например, у Dimetrodon, септомаксилла развита по всему заднему краю ноздри и достигает наверху носовой кости, образуя вдающийся в ноздрю передний «дерминтермедиальный» отросток. От верхнечелюстной кости — а у Haptodontinae также от слезной кости — септомаксилла отделена отверстиями. Квадратноскуловая кость укорочена, оттеснена от края височного окна и сближена с квадратной костью. Затылок наклонен вперед, то есть голова поднималась вверх из положения с растопыренными ногами. Затылочная поверхность высокая, широкая и вогнутая. Верхнезатылочная кость с боковыми отростками, проходящими над задневисочными окнами. Удлиненные околозатылочные отростки достигают чешуйчатой кости и отогнуты вниз и назад.
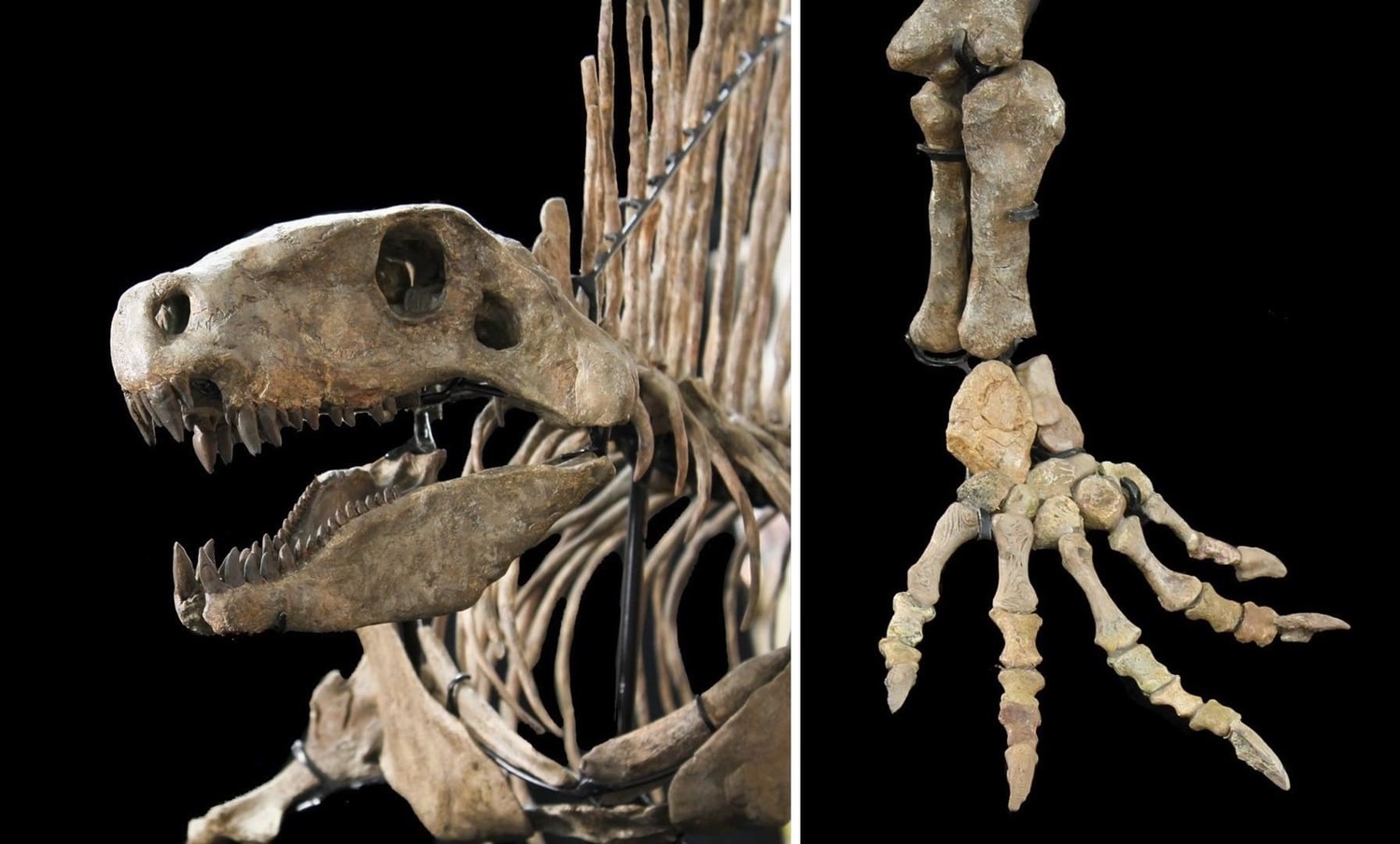
Смонтированный скелет продвинутого сфенакодонта Dimetrodon из ранней перми США.
Чтобы обеспечить опору зубному ряду, верхнечелюстная кость продвинутых сфенакодонтов разрастается далеко вверх в передней части черепа, отделяя слезную кость от наружной ноздри. Край предчелюстной кости у Sphenacodontinae сильновыпуклый. Предполагается, что у сфенакодонтов была крупная носовая полость и острое обоняние. Край верхней челюсти впереди клыков поднимается вверх в виде уступа или образует вырезку; судя по всему, эта черта связана со способом хватания добычи. Нижний край верхней челюсти сильно изогнут.
Небо глубоко вогнутое. У Sphenacodontinae из-за мощных верхних клыковидных зубов хоаны на их уровне сужены. Базиптеригоидные отростки короткие. Боковые отростки крыловидных костей массивные. Небные зубы никогда не покрывают парасфеноида. Поверхность квадратного мыщелка изогнутая. Челюстное сочленение сильно опущено по отношению к зубному ряду и поперечно расширено.
Череп с нижней челюстью Dimetrodon.
Нижняя челюсть высокая, с массивным симфизом. Сочленовная кость сильно увеличена, а ее суставная поверхность простирается по медиальной стороне нижней челюсти далеко вниз. Угловая кость с большой вырезкой.
Главный апоморфный признак сфенакодонтов — появление на угловой кости в задней части нижней челюсти округлой наружной отогнутой пластинки, или ангулярного лепестка, прикрывающего главную часть угловой кости. Функции этого лепестка неясны. У цинодонтов он участвует в формировании полости среднего уха маммального типа, но у сфенакодонтов, судя по всему, имел другое назначение. Предполагают, что их лепесток прикрывал заднюю крыловидную мышцу нижней челюсти, однако значение этого признака остается неясным. Согласно другой точке зрения, ангулярный лепесток все же представляет собой зачаток уха.
Прорисовки реконструированного черепа с нижней челюстью базального сфенакодонта гаптода (Haptodus betlei): череп сверху (А), снизу (Б) и сбоку (В), нижняя челюсть изнутри (Г).
Каким образом сфенакодонты воспринимали звуки, непонятно. У ранних синапсид (да и у прочих примитивных наземных позвоночных) органы слуха были плохо развиты. Важнейшая из слуховых косточек, стремя, входила у них в состав основания черепа. Но эта массивная кость вряд ли хорошо проводила звук. Большинство тетрапод того времени ощущали вибрации почвы, прижимаясь к земле головой и всем телом. Однако у сфенакодонтов вследствие удлиненности конечностей голова была приподнята над землей достаточно высоко. Высказано предположение, что ангулярный лепесток вибрировал и проводил звуки через угол нижней челюсти к стремени и ко внутреннему уху. Возможно, имелся и какой-то наружный слуховой проход позади задней стенки черепа. По крайней мере, у терапсид звукопроводящая система примерно такая же.
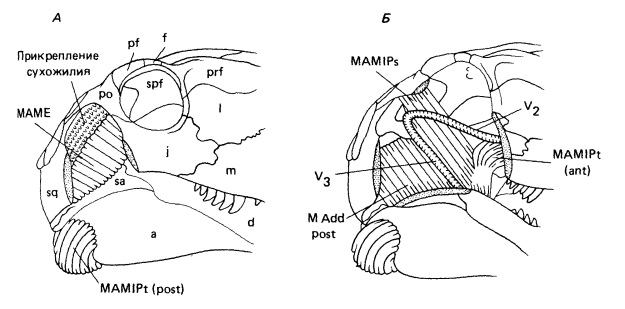
При сравнении с другими ранними амниотами изменения в конфигурации аддукторной камеры наводят на мысль об иной ориентации челюстных мышц. Вместо вертикального простирания от крыши черепа до дорсального края нижней челюсти они отклонялись вперед. Считают, что это могло обеспечить лучшее сопротивление вырывающейся добыче, способной сместить нижнюю челюсть вперед. Сходная по направлению сила, вероятно, возникала и при отрывании сфенакодонтами кусков мяса от крупной добычи.
Реконструкция челюстной мускулатуры прогрессивного сфенакодонта Dimetrodon: поверхностные (А) и глубинные (Б) слои. Антеровентральная ориентация основных мускулов-аддукторов, возможно, противодействовала направленным вперед силам, генерируемым бьющейся добычей. Сокращения: М Add post — задний аддуктор нижней челюсти, МАМЕ — наружный аддуктор нижней челюсти, MEMIPs — внутренний аддуктор нижней челюсти, ложновисочная мышца; MAMIPt (ant) — внутренний аддуктор нижней челюсти, крыловидная (передняя) мышца; MAMIPt (post) — внутренний аддуктор нижней челюсти, крыловидная (задняя) мышца; V2 — верхнечелюстная ветвь V-го нерва; V3 — нижнечелюстная ветвь V-го нерва.
Поскольку челюстной сустав находится гораздо ниже уровня зубного ряда, челюстные мышцы прикреплялись к наклонному дорсальному краю нижней челюсти, поднимающемуся с образованием выступающего венечного отростка. Разрастание этого отростка позволяет удлинить плечо рычага для смыкания челюстей, положение которого становится почти перпендикулярным к волокнам аддукторов.
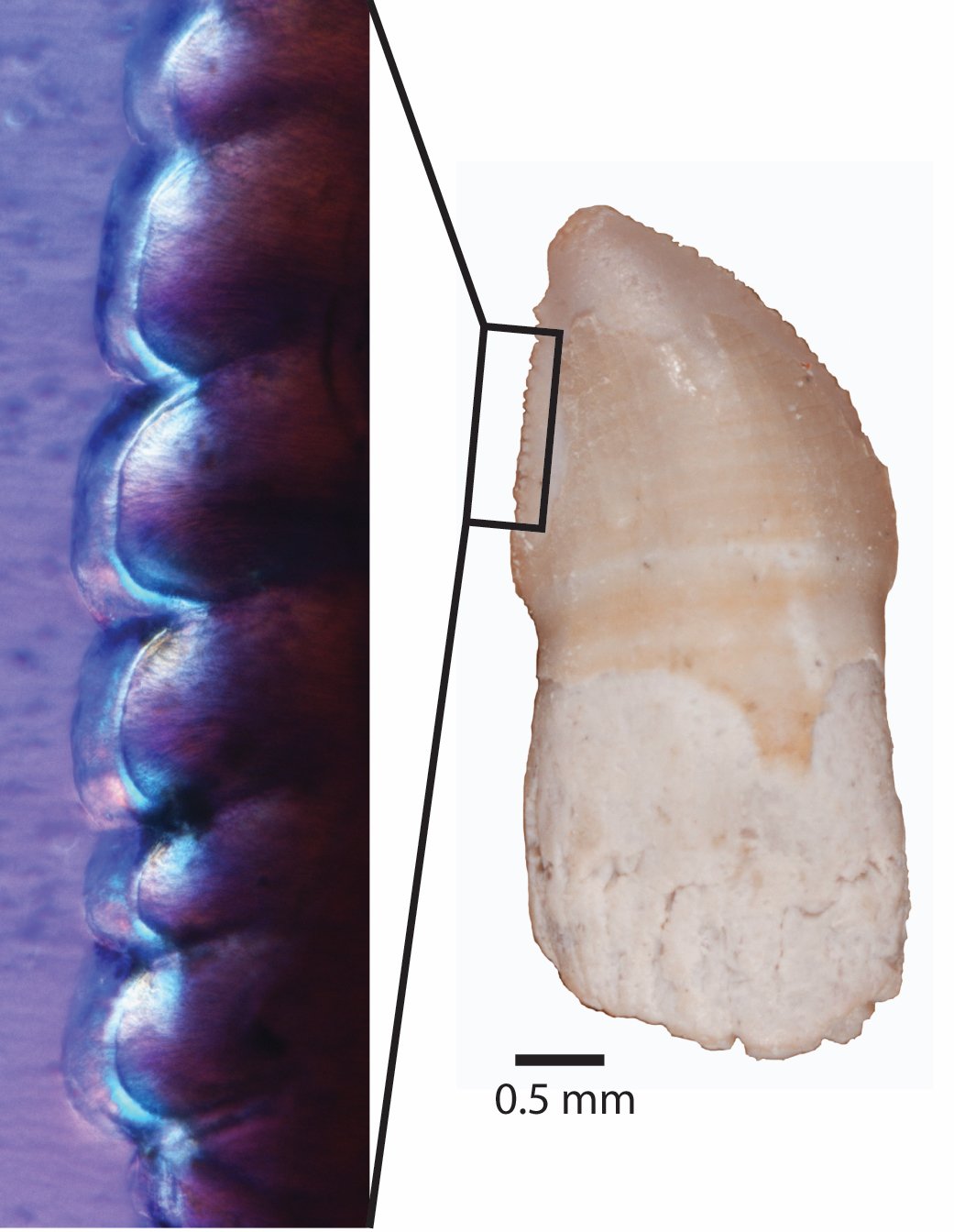
Зубы сидят в глубоких альвеолах, острые, пильчатые по передней и задней граням. Верхние и нижние клыковидные зубы развиты слабо (у Haptodontinae и Secodontosaurinae) или сильно (у Sphenacodontinae). В последнем случае челюстные зубы впереди клыковидных зубов редуцированы, и между предчелюстной и верхнечелюстной костями образуется глубокая вырезка для нижнечелюстного клыковидного зуба. Обычно с каждой стороны имеется 14-18 верхнечелюстных и 20-25 нижнечелюстных зубов.
Пильчатость челюстного зуба Dimetrodon.
Туловище средней длины, приземистое. Тело атланта достигает уровня вентральной поверхности позвоночника. Передние позвонки сжаты с боков и несут сильный вентральный киль. Туловищные позвонки умеренно или сильно удлиненные. В крестце 2-3 позвонка. Сочленовные площадки зигапофизов поставлены косо. Интерцентры умеренно развиты. Невральные дуги с вырезками над поперечными отростками; последние сильно удлинены и несколько смещены назад по сравнению с офиакодонтами. Остистые отростки позвонков удлинены незначительно (у Haptodontinae) или сильно (у Secodontosaurinae и Sphenacodontinae). Резко удлиненные отростки образуют на шее и спине высокий "парус". Остистые отростки прямые, без поперечных выростов, развитых у эдафозавров. Туловищные ребра двухголовчатые. Поясничные ребра укорочены.
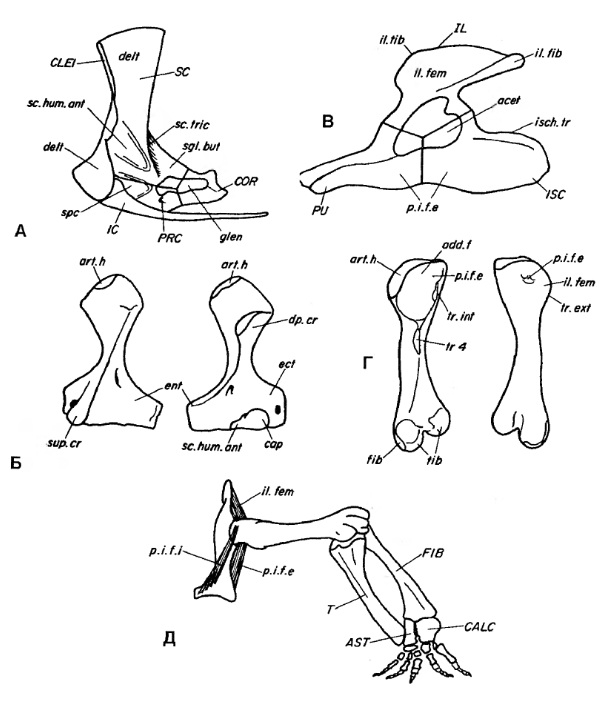
Вентральные отделы ключиц и тело межключицы расширенные. Подвздошная кость удлиненная, направленная назад и не суженная заметно над вертлужной впадиной. Конечности относительно длинные, передние и задние примерно равной длины. Аддукторный гребень и IV-й вертлуг бедренной кости слабо выражены. Кисть и стопа удлиненные. Гастралии развиты слабо. Хвост длинный.
Прорисовки скелета конечностей и их поясов прогрессивного сфенакодонта Dimetrodon: А, Б — передняя конечность; В, Г, Д — задняя конечность.
Сфенакодонты появились в середине позднего карбона Северной Америки и Европы, достигнув расцвета в ранней перми этих регионов. Кости большинства видов обнаружены в красноцветных толщах юга США, где богатая лесная растительность окаймляла лагуны мелководного моря. Сфенакодонты доминировали как наземные активные хищники, убивавшие и разделывающие крупную добычу с помощью своих колюще-режущих зубов. Судя по телесным пропорциям, они являлись довольно подвижными животными. Строение склеротикального кольца свидетельствует о ночной активности.

Диметродон (
Dimetrodon limbatus)
в естественном окружении. Автор — З. Буриан.
На какой-то стадии своей эволюции сфенакодонты дали начало терапсидам. Конкретные промежуточные формы между этими группами неизвестны, но все морфологические особенности указывают на существование таких форм. Возможно, они обитали в тех областях, где захоронение ископаемых остатков было практически невозможным (например, вдали от водоемов, на водоразделах), в то время как почти все пеликозавры населяли зону влажного тропического климата и берега водоемов.
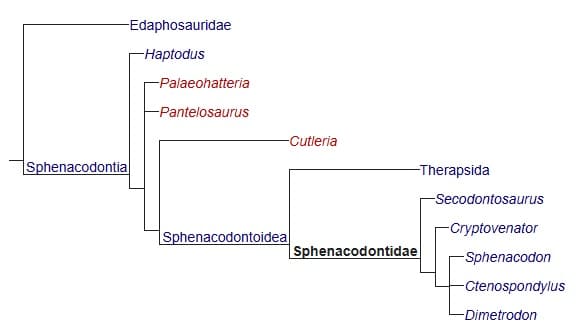
Кладограмма, отображающая родственные связи сфенакодонтов (по Фребишу с соавт., 2011 г.).
В современной кладистической систематике клада Sphenacodontia используется для обозначения монофилетичной группы, включающей сфенакодонтов и всех их потомков (включая млекопитающих). Подчиненная клада Sphenacodontoidea объединяет специализированных сфенакодонтов Sphenacodontidae (таких, как Secodontosaurus, Sphenacodon и Dimetrodon) и терапсид. Более ранние примитивные формы сфенакодонтов наподобие Haptodus, Palaeohatteria, Pantelosaurus и Cutleria (в докладистической классификации все включались в род Haptodus) выведены за рамки Sphenacodontoidea.
Ссылки
Sphenacodontia (Paleobiology Database)
перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Иванов А. О., Черепанов Г. О. Ископаемые высшие позвоночные. — СПб.: Издательство Санкт-Петербургского университета, 2001
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГЕОС, 2009