Синоконодоны — Sinoconodon
Клада Млекопитающеобразные (Mammaliaformes)
Отряд incertae sedis
Семейство †Синоконодонтовые (Sinoconodontidae)
Род †Синоконодоны (Sinoconodon)
Вид †Sinoconodon rigneyi
Синоконодоны (Sinoconodon Patterson et Olson, 1961) — монотипический род монотипического семейства синоконодонтовых (Sinoconodontidae Mills, 1971), происходящий из ранней юры Азии.
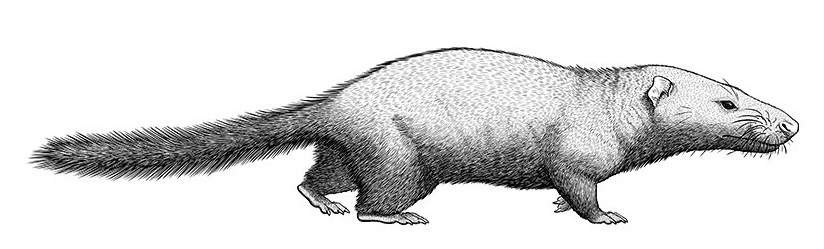
Реконструкция внешнего облика синоконодона (Sinoconodon rigneyi). Автор — Alphynix.
Синоним: Lufengoconodon Young, 1982.
Единственный установленный вид — Sinoconodon rigneyi Patterson et Olson, 1961 (тип). Его остатки обнаружены в провинции Юньнань, Китай. Они датируются ранней юрой (нижние слои формации Люфенг, синемюрский ярус). Синонимы: Lufengoconodon changchiawaensis Young, 1982; Sinoconodon changchiawaensis Crompton et Sun, 1985; Sinoconodon parringtoni Young, 1982; Sinoconodon youngi Zhang, 1983.
Родовое наименование в переводе с древнегреческого языка означает "китайский конический зуб".
Череп
Обилие синонимов рассматриваемого таксона обусловлен тем, что найденные остатки принадлежат особям различного возраста: наименьший известный череп составляет приблизительно 2 см в длину, тогда как самый крупный достигает свыше 6 см в длину и 2,3 см в ширину. Такой череп соответствует общей длине тела в приблизительно 35 см. Вес мог достигать 500 г.
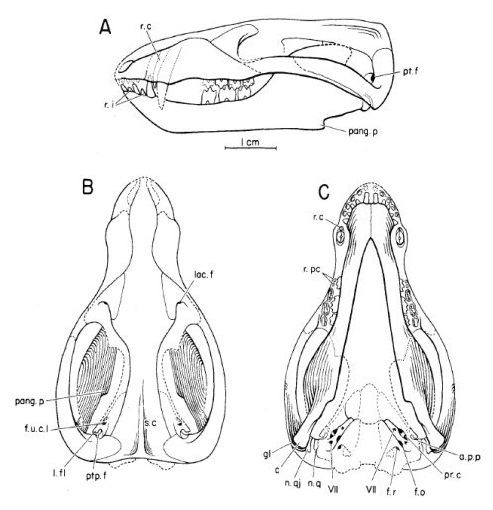
Прорисовки реконструированного черепа с нижней челюстью синоконодона (Sinoconodon rigneyi): сбоку (А), сверху (В) и снизу (С).
Голова большая относительно тела. Предчелюстная кость имеет выступ, расположенный посредине между двумя носовыми костями. У его небного отростка наличествует углубление для вхождения кончика нижнего клыка. Крупная септомаксилла выглядит формирует боковые границы наружных ноздрей. У нее имеется поперечный выступ, накладывающийся на носовую сторону предчелюстной кости, с септомаксиллярным отверстием в шве септомаксиллярно-челюстного соединения. Верхняя челюсть имеет 2 подглазничных отверстия и третье в челюстно-скуловом шве. Слезные кости большие, с обширной лицевой частью.
Глазница крупная. Среднеглазничная стенка укреплена выростами глазоклиновидной кости и глазничным выростом небных костей. Алисфеноид в виде широкой ивертикальной пластинки. Предшествующая тонкая пластинка верхнего края височной кости широкая, с 2 отверстиями для нижнечелюстных ветвей тройничного нерва и отверстия в алисфеноидно-височном шве для его верхнечелюстной ветви. Выпуклый передний околозатылочный отросток подобен таковому Tritylodontidae; снаружи он прикрыт чешуйчатой костью.
Передний и задний околозатылочные отростки разделены широким углублением, простирающимся от ямки мышцы стремени; данная ямка не слишком глубокая вентрально, что также характерно для Morganucodon и Dinnetherium. Боковой гребень височной кости перекрыт ветвью эпиптеригоида квадратной кости. Его передняя и вертикальная часть прободена 2 сосудистыми отверстиями. Боковая ложбинка височной кости демонстрирует открытый предушной канал, лицевой отверстие и, возможно, канал височного нерва. Подобно млекопитающим, имеется мысок височной кости. Он меньше, чем у Morganucodon, с плоской срединной поверхностью. Кроме того, предушная стенка мозгового черепа образована не алисфеноидом, как у сумчатых и плацентарных, а выростом периотики, как у однопроходных.
Тело клиновидной кости с двумя маленькими внутренними отверстиями для прохождения сонных артерий, но базисфенидное крыло отсутствует. Небные кости имеют большой глазничный выступ, формирующий заднюю стенку носовой впадины. Небные кости значительно больше, чем у цинодонтов и формируют края глоточного прохода, смещая птеригоидный крючковидный отросток назад. У костной крышки зева наличествует срединный гребень и 2 крылонебных гребня. Крыловидный отросток выглядит маленьким в сравнении с цинодонтами и имеет на боковой поверхности поперечную мышечную ямку в форме полумесяца.
Челюстной сустав маммального типа, сформированный мыщелком зубной кости и впадиной чешуйчатой кости. Строение нижней челюсти и среднего уха в целом сходен с таковым Morganucodonta. Как у Morganucodonta и Docodonta, имеется постдентальная ложбинка (хотя и относительно небольшая по сравнению с Morganucodon), а также развитый ложноугловой выступ с угловой впадиной.
Для млекопитающих характерны две апоморфии ушной области: удлиненный улитковый канал и возвышенность на барабанной поверхности кохлеарного региона, называемого мыском верхнего края пирамиды височной кости. У цинодонтов улитка короче, меньше и сориентирована более медиально, чем улитковый канал млекопитающих; их кохлеарный регион сформирован предушной областью и базисфеноидом. Sinoconodon имеет полностью сформированный мысок верхнего края височной кости, но короткую улитку, которая расширяется лишь на 1/3 длины мыска.
Кохлеарный канал Morganucodon вдвое длиннее, чем у Sinoconodon как относительно черепа, так и мыска. Более поздние млекопитающеобразные — например, Haldanodon и Multituberculata — демонстрируют более длинный улитковый канал. Предполагается, что мысок, преобразованный расширением височной кости за счет базисфеноида (вероятно, базиокципитально) является маркером преобразования цинодонтов в млекопитающих. Это изменение в окостенении и ориентации кохлеарного региона эффективно для лучшей звуковой изоляции внутреннего уха.
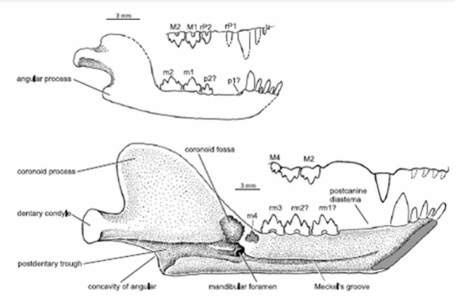
Прорисовки челюстей молодой (вверху) и взрослой (внизу) особей синоконодона (Sinoconodon rigneyi).
Зубы
Полная зубная формула: I5/i5, C1/c1, Pm2/pm2, M5/m5.
Подобно цинодонтам, резцы, клыки и задние щечные зубы на протяжении жизни неоднократно сменялись, постепенно увеличиваясь в размерах по мере роста животного. Число функционирующих резцов является переменным. На некоторых обнаруженных черепах с каждой стороны наличествуют лишь 3 резца, тогда как остальные находятся в процессе замены. Резцы имеют крупные и небольшие размеры. Попеременное расположение маленьких и больших зубов типично для цинодонтов, имевших их непрерывную и попеременную смену. Клыки сменялись, по меньшей мере, 4 раза. Размеры клыков сильно варьируют. Полностью развитые клыки имеют форму кинжала. Верхний клык настолько крупный, что может достигать нижнего края зубной кости, но на ранней стадии роста его коронка короткая и коническая.
Щечные зубы различаются по величине и имеют переменное число у разных особей. С возрастом предкоренные последовательно выпадали и уже не восстанавливались, не сохранившись ни на одном из найденных черепов. Это вело к образованию постепенно увеличивающейся заклыковой диастемы, сходной с таковой гомфодонтных цинодонтов. У молодых особей имеется по 5 верхних и нижних коренных зубов, однако во взрослом возрасте их число сокращается до 3. Передние 2 зуба утрачиваются без замены, что еще больше увеличивает заклыковую диастему. Данное явление также наблюдается, хотя и в меньшей степени, у Morganucodonta и возможно, у Hadrocodium и Kuehneotherium. Полная замена щечных зубов Sinoconodon, вероятно, напоминает таковую цинодонта Diademodon.
У молодых особей I-й верхний предкоренной конической формы, II-й — листовидный. Нижние щечные зубы в целом трехвершинные, однако демонстрирующие широкий диапазон морфологической изменчивости, возможно связанной с их заменой. Подобно цинодонтам, бугры на коренных зубах расположены строго линейно. На предпоследних коренных у некоторых крупных экземпляров имеется срединный цингулидный бугорок, отсутствующий на соответствующих зубах черепов поменьше. У верхнего коренного небольших экземпляров трехвершинная коронка с острыми выступами. В отличие от этого, верхний коренной больших экземпляров имеет тенденцию к развитию дополнительного дистального бугра. Вследствие вариативности размеров и строения верхних и нижних щечных зубов точного их смыкания не было, так что зоны износа на коронках отсутствуют.
Строение зубочелюстного аппарата Sinoconodon указывает на питание беспозвоночными и мелкими позвоночными животными.
Посткраниальный скелет
О посткраниальной анатомии Sinoconodon известно немного. Его плечевой пояс и передние конечности по своему строению мало отличаются от такового продвинутых цинодонтов — например, Bienotheroides. Как у однопроходных, присутствует большая межключица, неподвижно сочлененная с ключицей. Большой клювовидный отросток входит в состав ямки плечевого сочленения. У лопатки имеется акромион и надостная ямка на ее верхнем крае. Для плечевой кости характерна сферическая головка и большая бугристость, которая вместе с дельтопекторальным гребнем дистально продолжается приблизительно до половины длины плеча.
Из всех элементов таза и задней конечности известна лишь бедренная кость, характеризующаяся головкой, отделенной от большого вертлуга глубокой впадиной. Неполный малый вертлуг сохранился на средней стороне бедра. На его дистальном конце отсутствует пателларное углубление. Все эти особенности также имеются, хотя и в несколько отличном виде, у цинодонтов Tritheledontidae и Tritylodontidae, что делает их плезиоморфными для млекопитающих. К сожалению, имеющегося материала недостаточно для анализа особенностей локомоции Sinoconodon.
В отличие от других известных млекопитающеобразных, рост тела Sinoconodon, по-видимому, не прекращался в течение всей жизни. Судя по размерному диапазону найденных черепов, рост по мере взросления был постепенным, как у примитивных амниот, а не кратковременным и интенсивным, как это характерно для млекопитающих. Посткраниальных элементов для подтверждения этой точки зрения найдено не достаточно, однако есть основания предполагать, что увеличение общих размеров Sinoconodon коррелировало с многократной сменой его зубов.
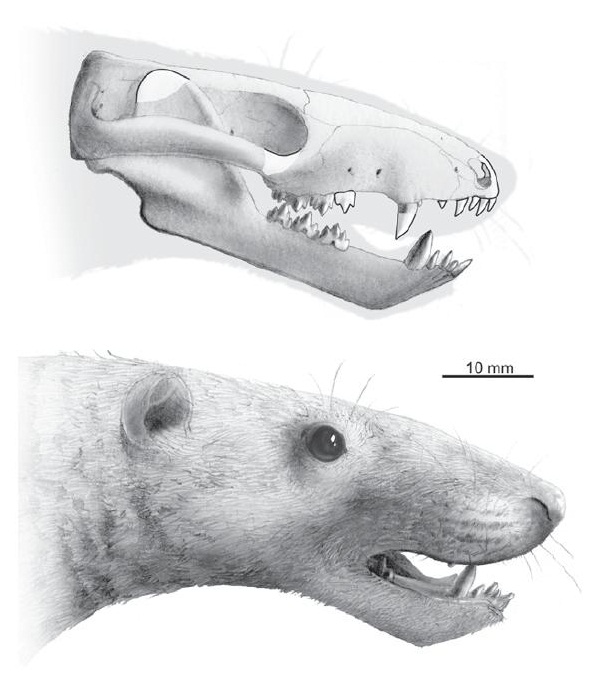
Прорисовка реконструированного черепа с нижней челюстью и портретная реконструкция синоконодона (Sinoconodon rigneyi). Автор — А. Hołda-Michalska.
Сравнение
Sinoconodon демонстрирует необычное сочетание примитивных и относительно продвинутых морфологических черт. Подобно цинодонтам, у него сохраняется большая септомаксилла, но в отличие от них височная кость имеет мысок. Кохлеарный регион устроен примитивнее, чем у Morganucodonta. Однако Sinoconodon напоминает Morganucodon строением каменистой кости: она также заходит ниже области полулунного ганглия, а ее передняя пластина окружает мандибулярную и максиллярную ветви тройничного нерва. Сочленовный мыщелок на зубной кости развит сильнее, чем у Hadrocodium, Morganucodonta и Kuehneotheriidae. Еще одно отличие от Morganucodonta, Docodonta и Kuehneotheriidae состоит в том, что меккелева ямка параллельна нижнему краю зубной кости, вместо того, чтобы сходиться на нем.
Если судить по размерам и глубине гленоидной впадины, а также степени редукции постдентальных костей, Sinoconodon более специализирован, чем Morganucodon. Челюстной сустав находится явно ниже уровня зубного ряда, как у хищных цинодонтов, в отличие от его более дорсального положения у Morganucodon. Щечные зубы, как и у последнего, латерально сжаты и несут продольный ряд бугров, но точная окклюзия отсутствует. Такое строение, несомненно, примитивно и, видимо, связано с прочным соединением ветвей нижней челюсти в симфизе, не позволяющем им двигаться независимо друг от друга и поворачиваться вдоль продольной оси.
Подобно цинодонтам и в отличие от типичных млекопитающих, зубы Sinoconodon в течение жизни неоднократно сменяются, за исключением навсегда утрачиваемых передних щечных зубов. Последнее иногда наблюдается и у Morganucodon, но у Sinoconodon передние щечные зубы не сохранились ни в одном из найденных черепов. В результате их незаменяемости между клыками и оставшимися щечными зубами образуется широкая диастема. Ее наличием Sinoconodon отличается от Morganucodonta, Kuehneotheriidae и Docodonta, но не от Hadrocodium. В отличие от Kuehneotheriidae сохраняется примитивное линейное расположение 3 основных бугров на щечных зубах.

Синоконодон (Sinoconodon rigneyi) в естественном окружении. Автор — К. Эулефи.
Систематика
Систематика
При первописании Паттерсон и Олсон (1961 г.) отнесли Sinoconodon к Triconodontidae вследствие трехбугорчатой формы коронок коренных зубов и некоторых особенностей черепного строения. В дальнейшем стало понятно, что эти морфологические черты характерны не только для Sinoconodon с Triconodontidae, на также распространяются на Triconodonta, Amphilestidae, Morganucodonta некоторых цинодонтов. Впоследствии Sinoconodon был отнесен к Morganucodontidae ("Eozostrodontidae") наряду с Morganucodon, Megazostrodon и Erythrotherium, основываясь главным образом на особенностях строения нижней челюсти и расположения зубов. Миллз (1971 г.) выделил семейство Sinoconodontidae, включающее Sinoconodon и Megazostrodon, базируясь на предположении, что Sinoconodon не имел блокировок коренных зубов, как это наблюдается у Megazostrodon.
Кромптон (1964, 1974 гг.) отметил, что Sinoconodon имеет несколько признаков нижнечелюстного строения, характерных для цинодонтов — в частности, отсутствует цингулид, имеющийся у Megazostrodon. Основываясь на этих соображениях, Кромптон (1974) исключил Megazostrodon из Sinoconodontidae, что впоследствии подтвердили Чжанг и Цуи (1983 г.). Кромптон и Сунь (1985 г.) отнесли Sinoconodon к млекопитающим, оговорив при этом, что он дивергировал от самого основания этой линии на более примитивном уровне, чем Morganucodonta. Поскольку о примитивных Adelobasileus и Haramiyida было мало что известно, Sinoconodon некоторое время принимался в качестве сестринской группы по отношению к остальным млекопитающим. Проанализировав морфологические особенности, Ло и Ву (1994 г.) показали, что Lufengoconodon changchiawaensis, Sinoconodon changchiawaensis, Sinoconodon parringtoni и Sinoconodon youngi являются младшими синонимами Sinoconodon rigneyi.
В целом рассматриваемый таксон считается более примитивным и обладает менее выраженными признаками млекопитающих, чем большинство других базальных млекопитающеобразных, включая Morganucodonta и Docodonta. Вероятно, он является реликтом более древней стадии эволюции млекопитающеобразных, либо представляет их рано уклонившуюся ветвь.
Ссылки
Синоконодон (Википедия) перейти
Sinoconodon (Paleobiology Database) перейти
Литература
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Татаринов Л. П. Эволюция териодонтов. — М.: Наука, 1976
Averianov A. O., Lopatin A. V., Skutschas P. P., Martynovich N. V., Leshchinskiy S. V., Rezvyi A. S., Krasnolutskii S. A., Fayngertz A. V. Discovery of Middle Jurassic mammals from Siberia. Acta Palaeontologica Polonica 50 (4), 2005
Kemp T. S. The Origin and Evolution of Mammals. — OUP Oxford, 2005
Kielan-Jaworowska Z., Cifelli R. L., Luo Z.-X. Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure. — Columbia University Press, 2004
Luo Z.-X., Kielan-Jaworowska Z., Cifelli R. L. In quest for a phylogeny of Mesozoic mammals. Acta Palaeontologica Polonica 47 (1), 2002
Luo Z.-X., Crompton A. W., Lucas S. G. Evolutionary origins of the mammalian promontorium and cochlea. Journal of Vertebrate Paleontology 15(1), 1995
Rose K. D. The beginning of the age of mammals. — JHU Press, 2006