Приветствую Вас Гость | RSS
Век млекопитающих - Age of Mammals
Меню сайтаФорма входа |
Систематика В 1895 г. Г. Сили выделил в составе высших синапсид (по его классификации — Therosuchia), кроме териодонтов и аномодонтов, или дицинодонтов, новую базальную группу диноцефалов (Dinocephalia). Эта классификация была фактически общепринятой до 1950-х гг. Однако открытие древних терапсид казанского и уржумского возраста в Приуралье привело к мысли о целесообразности внесения изменений в эту классификацию. В 1956 г. Д. Уотсон и А. Ромер разделили диноцефалов на хищных и растительноядных и включили первых в число териодонтов, а вторых — в число аномодонтов. Ю.А. Орлов (1958 г.) склонен был сближать хищных диноцефалов с териодонтами. Э. Олсон (1962 г.) установил инфраотряд Eotheriodontia, включающий фтинозухид, биармозухид (вместе с Eotitanosuchus) и бритоподид. Л. Бунстра (1963 г.) исключил бритоподид из этой группы, которую он назвал "Eotitanosuchia". Э. Олсон (1971 г.) выделил базальную группу Eotitanosuchidae и Phthinosuchidae, а прочих диноцефалов разделил на 3 группы: 1) Deuterosauridae и Tapinocephalidae; 2) Estemmenosuchidae; 3) Titanophoneidae с Brithopodidae и Titanosuchidae. Но П.К. Чудинов (1983 г.) оставил бритоподид в составе диноцефалов. Л.П. Татаринов (1974 г.) же принял предложенный Л. Бунстра объем эотериодонтов. К концу XX в. широкое распространение получило разделение терапсид на 3 основные группы: базальные диноцефалы и боковые ветви — преимущественно хищные териодонты и растительноядные дицинодонты. А. Ромер (1966 р.) объединяет диноцефалов и дицинодонтов, а также сравнительно небольшие растительноядные инфраотряды Venyukoviamorpha и Dromasauria в подотряд Anomodontia. Попытку капитальной реконструкции классификации пеликозавров и низших терапсид с широкой ревизией их родственных соотношений предпринял М.Ф. Ивахненко (2002-2003 гг.). Он предположил, что основной процесс в эволюции синапсид связан с оптимизацией челюстного аппарата, что прослеживается при изучении строения аддукторной области этих рептилий. М.Ф. Ивахненко выделил 4 типа строения: офиакодонтный, сфенакодонтный, эотериодонтный (примитивно-терапсидный) и эвтериодонтный. Офиакодонтная конструкция проиллюстрирована на примере Mesenosaurus. Стенка аддукторной полости даже при сильном развитии височного окна узкая и медиальная ее сторона ограничена структурами квадратной ветви небно-квадратного хряща — широким эпиптеригоидом и высокой квадратной костью. Между этими структурами и латеральной стенкой мозговой коробки по всей высоте черепа сохраняется эпиптериковая полость, отделяющая заднюю часть первичной верхней челюсти от мозговой коробки. Верхняя головка квадратной кости связана с дорсальным флангом чешуйчатой кости, образующим 2 отростка — узкий и короткий латеральный для заглазничной кости и широкий медиальный, подстилающий надвисочную и теменную кости. При сфенакодонтной конструкции эпиптериковая полость сохраняется только в нижней части, высота квадратной кости уменьшена за счет разрастания чешуйчатой кости между площадкой для верхней головки квадратной кости и дорсальным флангом, подстилающим теменной щит. Область tectum роsteius мозговой коробки образует широкую субвертикальную пластину, выходящую к чешуйчатой кости над квадратной. М.Ф. Ивахненко предположил, что у сфенакодонтов часть челюстных аддукторов начала крепиться к передне-верхней стенке мозговой коробки. Эотериодонтная конструкция, характерная для горгонопов, диноцефалов и аномодонтов, по существу, отличается от сфенакодонтной только пропорциями. Эта конструкция проиллюстрирована на примере Biarmosuchus и горгонопа Suchogorgon. Вследствие дальнейшего уменьшения высоты квадратной кости и увеличения высоты надквадратной области чешуйчатой кости эпиптериковая полость превращается в узкий канал над квадратной ветвью птеригоида. Tectum posterius и tectum synoticum срастаются в единую вертикальную пластину, к боковой поверхности perioticum широко прикрепляются челюстные аддукторы. Чешуйчатая кость крепится к периотике по сторонам посттемпорального отверстия — по латеральному флангу supraoccipitale в районе околозатылочного отростка (в примитивном виде, видимо, через остаток supratemporale). Квадратно-скуловая кость теряет контакт с чешуйчатой костью, образуется вырезка в задней части боковой поверхности черепа. Эвтериодонтная конструкция, проиллюстророванная на примере тероцефала Karenites, резко отличается от вышеописанных. Задний отросток заглазничной кости короток, а верхний край височного окна образован теменной костью. Сохраняется высокая эпиптериковая полость, но ее латеральная стенка в задней части образована не квадратной костью, а особым вентролатеральным отростком чешуйчатой кости, впервые описанным у тероцефала Regisaurus. Чешуйчатоя кость не подстилает теменную, а широко перекрывает ее сверху. Специальной площадки для причленения tectum posterius нет, периотика и чешуйчатая кость контактируют только в области мастоидного отростка. М.Ф. Ивахненко считает, что, помимо приблизительного сходства формы, между чешуйчатой костью эотериодонтов и эвтериодонтов мало общего. Это утверждение проиллюстрировано изображениями чешейчатой кости горгонопида Suchogorgon и тероцефала Karenites. Особое значение М.Ф. Ивахненко придал различиям в топографии чешуйчатой кости синапсид. На основании этих данных он разделил тероморф на 2 группы: первая включает параквадратобазальных синапсид (Ophiacodontia, Sphenacodontia), а также Eotheriodontia (Gorgonopia, Dinocephalia, Gorgodontia и Anomodontia); вторая объединяет сквамобазальных синапсид (Eutheriodontia). В первой группе особое положение занимают офиакодонты, у которых эпиптериковая полость высокая, отсутствует ангулярный карман внутри от ангулярного лепестка и присутствует контакт между скуловой и квадратно-скуловой костями. Изначально в первой группе в височной области имелись 2 кости — надвисочная и чешуйчатая, но после редукции supratemporale чешуйчатая, расположенная у пеликозавров на palatoquadratum, оказалась расположенной на ушной капсуле. Эдафозавры включаются в сфенакодонтную подгруппу, поскольку они имеют сфенакодонтную конструкцию с узким эпиптеригоидом, выходом челюстных аддукторов к верхней части мозговой коробки, хорошо развитый ангулярный лепесток и короткую квадратно-скуловую кость. Л.П. Татаринов (2009 г.) отмечает, что в действительности эдафозавры не имеют ангулярного лепестка с ангулярным карманом, а из известных в этом отношении офиакодонтов широкий эпиптеригоид имеется только у Mesenosaurus и Tetraceratops. Но систематическое положение обоих родов не вызывает особых сомнений. В любом случае, горгонопов М.Ф. Ивахненко вывел из состава "настоящих" сквамобазальных териодонтов. М.Ф. Ивахненко (2005 г.) пришел к выводу об отсутствии гомологии чешуйчатой кости у параквадратобазальных и сквамобазальных синапсид. Автор считает несомненной гомологию чешуйчатой кости млекопитающих (и сквамобазальных эвтериодонтов) с supratemporale примитивных офиакодонтов и сфенакодонтов. "Чешуйчатая" кость параквадратобазальных синапсид, по М.Ф. Ивахненко, гомологична второй, ниже расположенной, кости височной области. Рудимент supratemporale М.Ф. Ивахненко обнаружил у горгонопида Suchogorgon. Сквамобазальных териодонтов, или эвтериодонтов, М.Ф. Ивахненко склонен выводить от предков, расположенных на сквамобазальном уровне. Однако Л.П. Татаринов (2009 г.) сомневается в правоте идеи М.Ф. Ивахненко относительно различной гомологии чешуйчатых костей у параквадратобазальных и сквамобазальных синапсид. 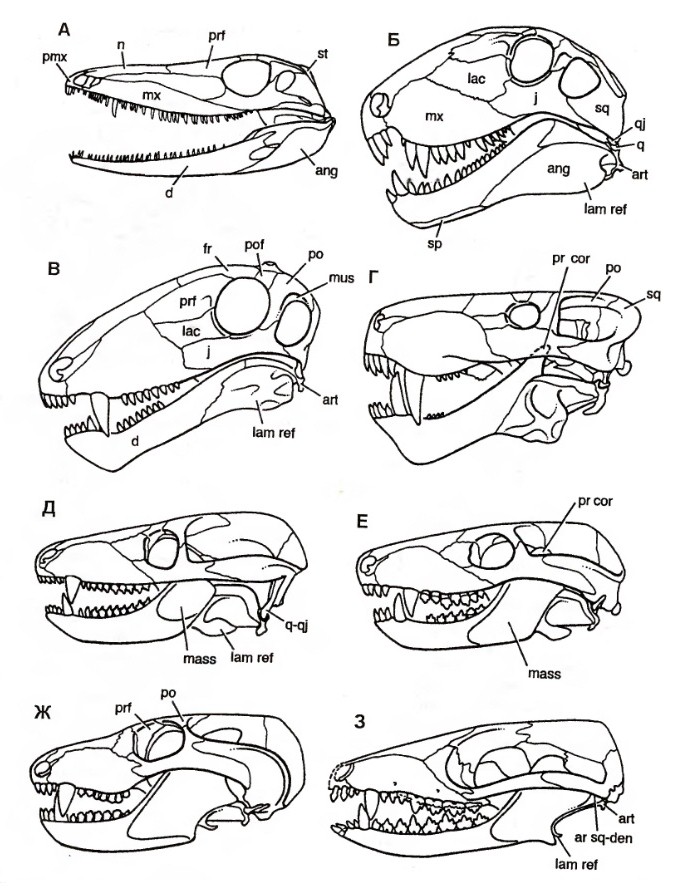 Прорисовки реконструированных черепов синапсид: A — пеликозавр Varanosaurus; Б — пеликозавр Dimetrodon; В — биармозухия Biarmosuchus; Г — горгоноп Leontocephalus; Д — цинодонт Procynosuchus; Е — цинодонт Thrinaxodon; Ж — цинодонт Probainognathus; З — примитивное млекопитающеобразное Morganucodon. Л.П. Татаринов не согласен с М.Ф. Ивахненко, считающим эпиптериковую полость эвтериодонтов (и млекопитающих) ограниченной снаружи отростком чешуйчатой кости. По мнению Л.П. Татаринова, это противоречит исходному определению эпиптериковой полости млекопитающих как пространства внутри от алисфеноида (эпиптеригоида, окостенения восходящего отростка исходного palatoquadratum) и вторично включенного в мозговую коробку у млекопитающих. На самом деле упомянутый отросток чешуйчатой кости расположен позади эпиптериковой полости и алисфеноида, хотя у млекопитающих он может разрастаться под теменной костью далеко вперед. У цинодонтов, у которых эпиптеригоид превращается в алисфеноид, он образует, по крайней мере, часть предушной стенки мозговой коробки. Сверху и сзади в образование предушной стенки у примитивных млекопитающих, в частности у однопроходных, может вносить свою лепту окостенение сфенообтураторной мембраны, связанной позади с ушной капсулой. Эта кость получила название запирательной пластины. 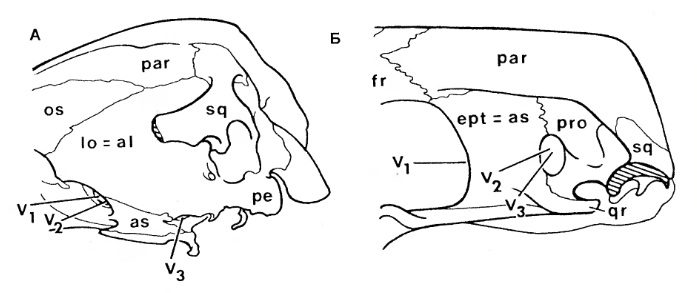 Прорисовки задней части черепа: А — прогрессивный цинодонт Pachygenelus; Б — однопроходное млекопитающее Ornithorhynchus. В группу "настоящих" териодонтов (эвтериодонтов) входят, как правило, хищные терапсиды с хорошо развитыми и сжатыми с боков клыками. Только у тритилодонтов, вторично ставших растительноядными, клыки отсутствуют. Не дифференцированы клыки и у отдельных тероцефалов, а также у проблематичных терапсид — Niaftasuchus и Nikkasaurus из уржумского яруса средней перми. Резцы конические, изредка долотовидные. Заклыковые зубы всегда однорядные, чаще конические, у многих тероцефалов и примитивных цинодонтов трехзубчатые, у бауриаморфов, гомфодонтных цинодонтов и у тритилодонтов поперечно расширенные, со сложным многорядным набором бугорков. Базикраниальная ось прямая, в передней части не изогнутая вниз, как у дицинодонтов. Затылок широкий, зачастую вогнутый с боков, почти вертикальный или верхней частью наклоненный вперед. Обе ушные кости обычно сливаются в периотику. Сошник почти всегда непарный, обычно без зубов. Крыловидные кости высших цинодонтов с хорошо развитыми поперечными отростками. У прогрессивных териодонтов за счет соединения небных отростков верхнечелюстных костей и, реже, небных костей образуется вторичное костное небо. Челюстное сочленение обычно двухмыщелковое, расположено впереди затылочного. Венечный отросток нижней челюсти обычно хорошо развит, у высших цинодонтов и бауриаморф он очень массивный. Независимо он образовался и у горгонопов. Традиционно териодонты считались монофилетической группой. Первоначально в их состав включили только исходно хищных терапсид с развитым венечным отростком, поднимающимся над верхним краем нижней челюсти. В таком объеме териодонтов разделили на горгонопов, тероцефалов, цинодонтов и вторично растительноядных гомфодонтных цинодонтов, и тритилодонтов. Но позднее исследователи выявили, что примитивные горгонопы — бурнетиоиды (Burnetioidea), а также отдельные тероцефалы — не имеют на нижней челюсти четко выраженного венечного отростка. Совершенно не развит венечный отросток и у примитивных эотериодонтов, исключая типичных горгонопов. Дальнейшее развитие системе терапсид придал М.Ф. Ивахненко (2003 г.), разделивший синапсид на сфенакодонтов, эдафозавров и собственно териодонтов. Последние разделены на Eotherapsida, или эотериодонтов (горгонопов, диноцефалов и аномодонтов), и Eutherapsida (собственно териодонтов) с мозговой коробкой алисфеноидного типа, с высокой и широкой эпиптериковой полостью. У Eotherapsida эпиптериковая полость редуцирована. Первая группа, таким образом, из числа терапсид объединяет диноцефалов, горгонопов и аномодонтов. Большая часть пеликозавров, кроме офиакодонтов, включена в сфенакодонтно-эотериодонтную группу. Формальный диагноз этих групп основан на отсутствии у первой из них высокой эпиптериковой полости, образованной медиальной стенкой аддукторной ямы между латеральной стенкой мозговой коробки (передней пластиной переднеушной кости) и единой вертикальной пластиной, образованной срастающимися друг с другом tectum posterius, tectum synoticum и opisthoticum. Чешуйчатая кость в этой группе контактирует с задним отростком заглазничной кости и подстилает теменную кость. У эвтериодонтной группы эпиптериковая полость высокая, медиальную стенку аддукторной ямы образуют расширенный эпиптеригоид и особый передний фланг чешуйчатой кости — "anteroventral process", отсутствующий у предыдущей группы. Чешуйчатая кость не контактирует с заглазничной и перекрывает теменную кость сверху. Л.П. Татаринов (2009 г.) считае преждевременным разделять всех синапсид на параквадратобазальных и сквамобазальных. Он сохраняет в традиционном объеме пеликозавров, рассматривая в их числе сфенакодонтов, а также таксон терапсид, в котором рассматривает как параквадратобазальных, так и сквамобазальных высших синапсид. Согласно Л.П. Татаринову, по принципам кладистической систематики можно, и даже удобнее объединить хотя бы сфенакодонтов с настоящими терапсидами. Принимаемую М.Ф. Ивахненко возможность разделения синапсид по размерам эпиптериковой полости (не редуцированной у эвтериодонтов и превращенной в узкий канал у эотериодонтов), Л.П. Татаринов не считаю строго доказанной. Во всяком случае, у немногих диноцефалов, у которых эпиптеригоид хорошо известен, его восходящая ветвь столь же высока, хотя и узка, как у примитивных цинодонтов и даже современных лепидозавров. У южноафриканского диноцефала Anteosaurus верхнюю часть восходящей ветви эпиптеригоида продолжает кзади необычно развитый сфенэтмоид, достигающий области шва между лобной и теменной костями, но ее отношение к эпиптериковой полости осталось неясным. Задний край восходящей ветви ограничивает отверстие для выхода тройничного нерва. Уже упоминавшийся внутренний отросток чешуйчатой кости выстилает нижний фланг теменной кости и проходит над ушной капсулой. Я считаю сомнительным, что отросток этот в прямом смысле слова выстилает эпиптериковую полость, хотя нужно отметить, что у Anteosaurus нижневнутренний его край граничит с внутренним краем самого верхнего конца восходящего отростка эпиптеригоида.  Парасагиттальный разрез черепа диноцефала антеозавра (Anteosaurus magnificus). Такая же узкая, но почти столь же высокая восходящая ветвь эпиптеригоида известна у Ulemosaurus из уржумского яруса Приуралья, у которого связи этой кости со сфенэтмоидом и чешуйчатой костью остались невыясненными. У Ulemosaurus эта ветвь имеет большое сходство с эпиптеригоидом лепидозавров. Узкий, но высокий эпиптеригоид, почти достигающий наверху области теменного отверстия, имеется у многих горгонопов, например у Gorgonops; правда, в работах эпиптеригоид горгонопов изображается как передняя предушная часть проотики, отделенная швом от основной части этой кости. Палочковидный умеренно высокий эпиптеригоид, по-видимому, имеется у примитивного цинодонта Dvinia, ошибочно описанный в качестве прегипофизарного отростка базисфеноида. У прогрессивных цинодонтов эпиптериковая полость в связи с ее вхождением в стенку эндокрания может быть очень небольшой, хотя это обстоятельство маскируется развитием в верхней части этой области запирательной пластины. Крошечную эпиптериковую полость обычно имеют и млекопитающие, у которых через нее проходит I-я (r. ophthalmicus profundus), а иногда II-я ветви (r. maxillaris) тройничного нерва. Но высокий и очень узкий восходящий отросток эпиптеригоида известен у ряда дицинодонтов хорошей сохранности — Dicynodontoides, Robertia и др. В целом, картина восходящего отростка эпиптеригоида, с точки зрения Л.П. Татаринова, не дает оснований для определения четких различий по этому признаку только 2 групп терапсид. 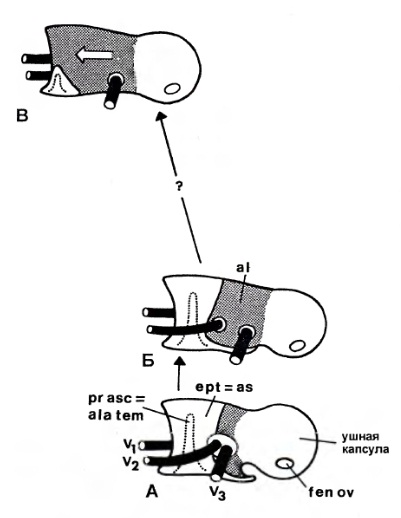 Схема строения задней части эндокрания: А — цинодонт; Б — раннеюрское млекопитающеобразное Morganucodon; В — однопроходное Ornithorhynchus. На сегодняшний день зарубежные исследователи чаще всего принимают терапсид как отряд класса синапсид (по другим классификациям — инфракласс или даже подкласс). В этот отряд обычно зачисляют такие основные группы: биармозухии (Biarmosuchia), диноцефалы (Dinocephalia), аномодонты (Anomodontia), а также териодонты, или зверозубые (Theriodontia), включающие горгонопсий (Gorgonopsia), тероцефалов (Therocephalia) и цинодонтов (Cynodontia). На приводимой ниже кладограмме представлены филогенетические связи между данными группами согласно распространенным ныне воззрениям: 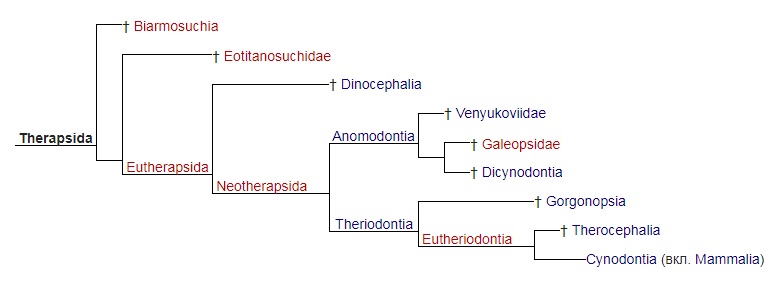 Поскольку клада, образуемая подотрядом цинодонтов (Cynodontia), включает также и класс млекопитающих, данный подотряд (а потому и отряд Therapsida в целом) является парафилетической группой. |
Поиск |