Приветствую Вас Гость | RSS
Век млекопитающих - Age of Mammals
Меню сайтаФорма входа |
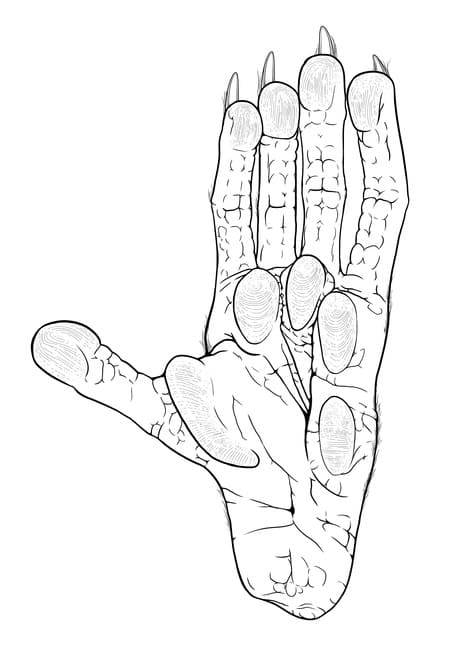
История изучения Впервые соневидный опоссум был описан Ф. Филиппи в 1899 г. в качестве Didelphis australis. Родовое название Didelphis указывает на принадлежность к опоссумам, а видовое australis указывает на южное распространение животного. Однако оказалось, что это название еще в 1812 г. было преоккупировано Гольдфуссом для другого сумчатого. Уже в 1894 г. Олдфилд Томас присвоил соневидному опоссуму валидное наименование — Dromiciops gliroides. Новое родовое название Dromiciops происходит от древнегреческого слова δρομικός ("резвый") и основано на сходстве соневидного опоссума с толстохвостым поссумом (Cercartetus nanus) из Австралии, одним из синонимов которого является Dromicia nana. Видовой эхпитет gliroides представляет собой комбинацию латинских glis, gliris ("соня", в более общем смысле "грызун") и греческой приставки -oides ("похожий на"). В 1982 г. Маршалл объединил оба названия и получил рекомбинированное Dromiciops australis, затем на протяжении нескольких десятилетий применявшееся в отношении соневидного опоссума. В своей работе «Млекопитающие Чили» за 1943 г. Уилфред Хадсон Осгуд установил 2 подвида соневидного опоссума — Dromiciops australis australis и Dromiciops australis gliroides, которые сейчас ввиду отсутствия четких отличительных признаков считаются сомнительными. Дальнейшие исследования показали, что таксон Dromiciops gliroides имеет глубокую филогеографическую структуру с 3 алопатрическими и высокодифференцированными группами, включая 2 непризнанных безымянных вида. Основываясь на морфологических различиях, выраженных главным образом в строении черепа и зубов, а также молекулярных и географических данных, в 2016 г. Д'Элия, Уртадо и Д'Анатро выделии из монотипического до этого Dromiciops gliroides 2 новых вида — Dromiciops bozinovici и Dromiciops mondaca. Видовой эпитет первому из них присвоен в честь териолога Франциско Божиновича, а второму — в честь Фреди Мондаки, смотрителя коллекции млекопитающих при Южном чилийском университете. Полагают, что род Dromiciops произошел от представителей рода Microbiotherium, известного из раннего миоцена Южной Америки, при этом некоторые авторы считают эти роды неотличимыми. Все остальные роды семейства Microbiotheriidae, такие как Pachybiotherium, вымерли к концу миоцена. Строение и функции Размеры мелкие: длина головы и туловища составляет 8,3-13 см, хвоста — 9-13,2 см, т. е. они лишь немного отличаются по длине. Общая длина варьирует от 19,5 до 24,6 см, масса тела — от 16 до 49 г. Половой диморфизм в линейных размерах практически не выражен, но самки перед зимовкой набирают относительно больший вес. Пропорциями зверьки напоминают мелких представителей семейства опоссумовых (Didelphidae). Голова с короткой заостренной мордочкой, крупными глазами и относительно небольшими (1-2 см в высоту), овальными, приподнятыми вверх ушными раковинами. Хрящевой луч, формирующий передний край раковины, мало развит, а сзади она покрыта короткими и густыми волосами, в отличие от практически голых ушей других сумчатых Западного полушария. На мордочке и вокруг глаз размещаются многочисленные вибриссы. В области ноздрей развит ринариум. Обыкновенный соневидный опоссум (Dromiciops gliroides) с жировыми отложениями в хвосте и без них. Кости конечностей относительно короткие и широкие, соединенные крепкими суставами. Кисть и стопа пропорционально больше и массивнее, чем у опоссумовых того же размерного класса. Длина кисти составляет 0,9-1 см, длина стопы — 1,5-2 см. Ступни пятипалые, с цепкими когтями. На задней конечности IV-й палец является самым длинным, II-й, III-й и V-й пальцы уступают ему по длине, а I-й палец наиболее короткий. I-е пальцы на передней и задней конечностях противопоставляются остальным, что способствует лучшему захватыванию веток. На подошве задней лапы имеется 5 поперечно исчерченных подушечек — 1 у основания I-го пальца, 3 у оснований II-IV-го пальцев и еще 1, самая маленькая, у наружной границы ступни. На концевых фалангах пальцев расположены крупные подушечки, имеющие продольную исчерченность и выступающие за пределы когтей. Хвост, толстый у основания, быстро и равномерно утоньшается к кончику. Он цепкий лишь в сравнительно небольшой степени. В отличие от опоссумовых, хвост густо покрыт волосами, кроме голого участка протяженностью 2,5-3 см и небольшой ширины на его нижней поверхности вблизи кончика, что улучшает сцепление с субстратом. Присутствие шерсти на хвосте, вероятно, имеет вторичный, а не изначальный характер и, возможно, коррелирует с уменьшением степени его цепкости. Передняя 1/3 хвоста служит для запасания жира и может вдвое увеличиваться в диаметре. На протяжении недели перед впадением в спячку за счет накопления жировых отложений зверьки способны удвоить свой вес.
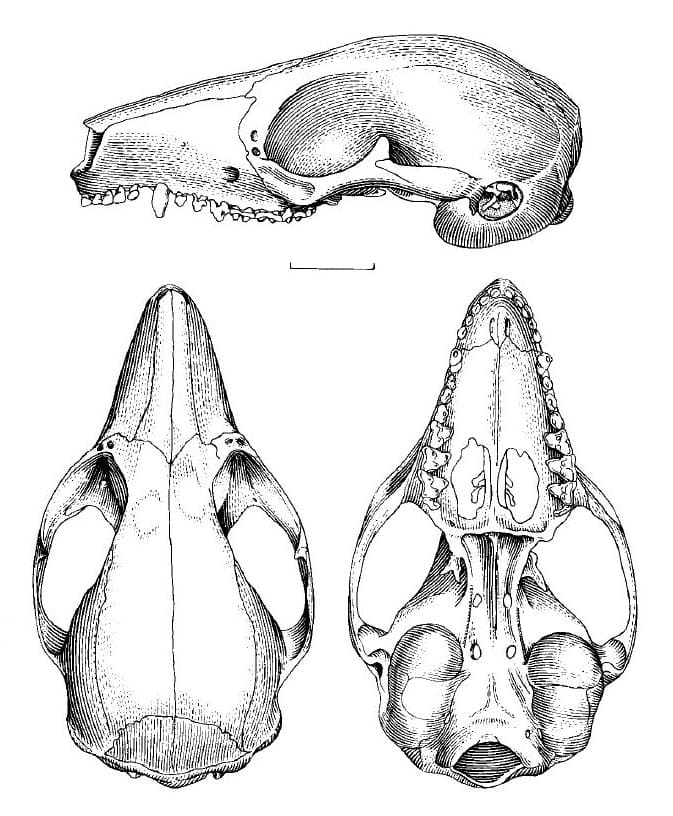
Меховой покров короткий, густой и мягкий. На верхней части тела и передней 1/3 хвоста его окраска довольно яркая, светло-коричнево-серая, причем спина явно темнее боков. Морда светло-серая, с четко выраженными черными кольцами вокруг глаз, иногда доходящими до носа, и более светлыми щеками. Над глазами и под ушами имеются слабо выраженные потемнения. Верхняя часть головы и шеи красновато-коричневые. Спереди и сзади плечей и перед бедрами расположены большие светло-серые пятна, лишенные отчетливых границ. На наружных поверхностях конечностей имеются темно-коричневые пятна, постепенно светлеющие к их дистальным отделам. Выше плечей пятна плавно переходят в темную спину, тогда как над бедрами они отграничиваются узкими светлыми областями, проходящими примерно вдоль тазовых костей. Тыльные стороны кистей и ступней покрыты редким светлым мехом, собранным на концевых фалангах в подобие кисточек. Цвет горла, брюха и внутренних поверхностей конечностей варьирует от светло-серого до желтовато-белого. Последние 2/3 хвоста покрыты более жесткими, чем на остальном теле, темно-коричневыми волосами. Окраска особей даже в одной популяции может сильно варьировать. Специфические кожные железы выделяют секрет с резким неприятным запахом, который может служить для отпугивания хищников или привлечения половых партнеров. У самок имеется небольшая, но хорошо сформированная выводковая сумка, открывающаяся вентрально и опушенная светло-коричневым мехом. В сумке находятся 4 соска, симметрично расположенные 2 парами. Медиальное псевдо-влагалище хорошо развито. У самца мошонка расположена на коротком черешке и также покрыта светло-коричневыми волосками. В отличие от всех других современных сумчатых и подобно однопроходным, клоака базикаудальная — т. е., находится в основании хвоста. По некоторым данным, она развита лучше, чем у прочих современных сумчатых. Череп с укороченным лицевым и округлым мозговым отделом. Предчелюстные кости сильно удлинены, что выражается в значительном развитии предклыковой части морды. Носовые кости расширяются кзади. Межглазничная область широкая. Отсутствуют какие-либо следы заглазничных отростков, височных линий и стреловидного гребня. Околозатылочных отростков также нет. Небо максимально расширено на уровне переднего края M3. Хоаны широкие, их мезиальная перегородка продолжается в виде узкого гребешка на основании черепа. Засуставный отросток чешуйчатой кости маленький, очень короткий и спереди слегка вогнутый. В отличие от опоссумовых, заднее сонное отверстие отсутствует. По результатам измерений большой выбоки, кондилобазальная длина черепа составляет 2,36-2,79 см, скуловая ширина — 1,44-1,68 см, ширина в области сосцевидных отростков — 1,15-1,26 см, межглазничная ширина — 0,45-0,53 см. У одного из черепов кондилобазальная длина составляет 2,6 см, скуловая ширина — 1,44-1,68 см, длина носовых костей — 1,1 см, их наименьшая ширина — 0,19 см, их наибольшая ширина — 0,44 см, длина неба — 1,4 см, его ширина на уровне переднего края M3 — 1 см, длина М1-3 — 0,46 см, длина m1-4 — 0,58 см. Слуховая область характеризуется целым рядом уникальных черт. Слуховой барабан по сравнению с другими сумчатыми чрезвычайно вздут и занимает значительную площадь в основании черепа. Барабан полностью окостеневает. Его передняя 1/3 составлена барабанным крылом алисфеноида (основной крылоклиновидной кости), образующим также переднюю восходящую часть внутреннего слухового прохода. Задние 2/3 барабана сформированы барабанным крылом каменистой кости, сильно пневматизированной сосцевидной костью, узкой пластиной базисфеноида, а также сильно развитой вентромедиальной костью между алисфеноидом и крыльями каменистой кости, определяемой в качестве внутренней барабанной кости. Эта кость отсутствует у других ныне живущих сумчатых, но широко распространена у плацентарных; ее форма и ориентация у микробиотериев уникальны и не встречаются у других млекопитающих. Она наличествует как самостоятельный элемент, отделенный швами от алисфеноида, каменистой кости и базисфеноида. Судя по всему, внутренняя барабанная кость не гомологична таковой у плацентарных. Вероятно, в данном случае эта кость является сильно вздутым производным каменистой части барабана, либо представляет собой придаточный элемент, который при развитии трехкомпонентного барабана заполняет медиовентральный и медиальный промежутки до того, как они закрываются алисфеноидом и каменистой костью, как это происходит у других сумчатых. Сбоку внутренняя барабанная кость формирует вентральную и каудальную части внутреннего слухового прохода. Медиально и медио-каудально она находится в шовном контакте с основной затылочной костью, далее каудально граничит с боковой затылочной костью и дорсально-каудально контактирует с небольшой сосцевидной костью. Таким образом, внутренняя барабанная кость полностью закрывает заднюю часть слухового барабана. Внешняя барабанная кость прикрывает вентральную поверхность слухового барабана, образуя небольшую выемку.
Примерно посередине слухового барабана, на его вентральной стороне, находится область овальной формы, граничащая спереди с вышеупомянутым алисфеноид-энтотимпаническим швом. Сзади она граничит с линией, соответствующей почти полной перегородке на внутренней стороне барабана. Барабанная полость разделена 2 перегородками на 3 отсека. Передняя перегородка, которая следует за швом между алисфеноидом и внутренней барабанной костью, низкая и имеет прямой дорсальный край. Она простирается от внутренней части латеральной стенки под барабаном, с которой контактирует, по полу барабана к ростральному концу улитки. Таким образом, передний отсек находится в широком контакте со средним, медиальная стенка которого сформирована околоушной костью, а латеральная — барабанной костью. Околоушная кость заканчивается спереди выпуклостью, которая отделена вентрально неглубоким пазом в медиальной части, проходящей дальше спереди и латерально. Вторая перегородка, которая отделяет средний отсек от заднего, лежит под каудальной частью барабанного кольца и соединяет его с дорсальной стороной вентральной стенки барабана. Перегородка в ее латеральной части проходит кзади от алисфеноидного шва, затем поворачивает медиально и принимает латеро-медиальное направление. Эта перегородка доходит до крыши полости, таким образом, полностью разделяя задний отсек посередине, за исключением небольшого отверстия овальной формы в его верхней части. Барабанная кость образует полное кольцо для расположенной дорсально маленькой барабанной вырезки. Данное пространство охватывает головку молоточка и тело наковальни. Эти элементы достигают эпитимпанальной выемки. Последняя отделена от наружного канала барабанной перепонкой и формируется в основном околоушной костью. Лишь латеральная стенка образована чешуйчатой костью. Костная пластинка возникает из дорсальной и задней подпорок барабанной кости и упирается в околоушную кость. Барабанная кость при взгляде сбоку не видна. Ось, на которой находятся слуховые косточки, расположена под углом от -5 до 10 ° по отношению к горизонтальной плоскости. Характерные особенности строения выражены у молоточка в гораздо большей степени, чем у 2 других косточек. Головка молоточка, если смотреть на нее сверху, имеет форму вытянутого овала с наиболее длинным диаметром практически в горизонтальной плоскости. В каудальном направлении суставные поверхности для причленения наковальни образуют угол около 90 ° по отношению друг к другу. Латерально на нижней границе головки молоточка расположен короткий изогнутый гребень. Он тянется каудо-вентрально с нижней стороны переднего конца головки до шейки молоточка и нависает над слегка рифленой проксимальной частью переднего отростка. Ось головки образует медиально открытый угол примерно 150 ° по отношению к шейке. Эта морфологическая особенность является уникальной среди сумчатых. Передний отросток, имеющий сложное строение у других сумчатых, в данном случае мал и упрощен. Его барабанная пластинка слита с барабанной костью и легко ломается при удалении молоточка. Рукоятка молоточка длинная; образуемый ею рычаг относительно больше, чем у других сумчатых. Твердое тело наковальни имеет хорошо развитую короткую подпорку, которая достигает наковальной ямки в задней стенке барабанной полости. Стремячко треугольной формы. Стремячковая пластинка имеет эллиптическую форму.
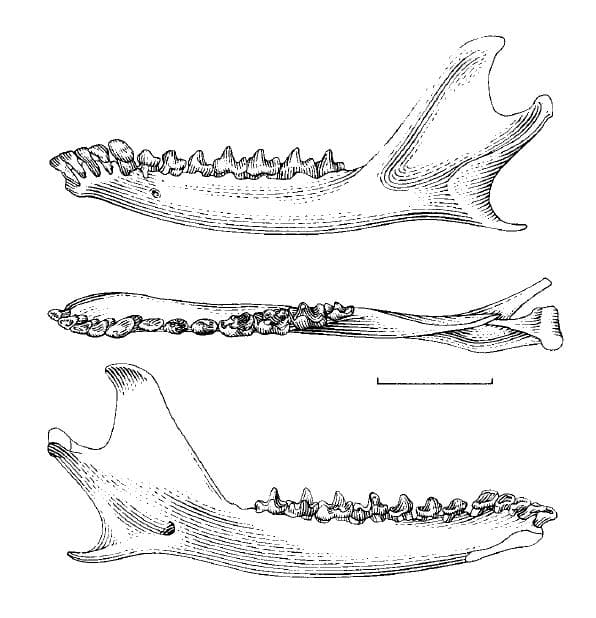
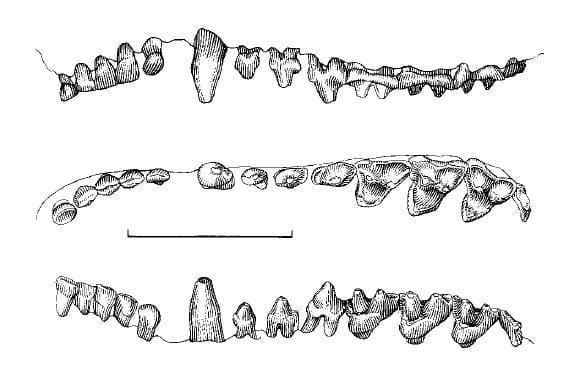
Нижняя челюсть с очень коротким симфизом, передняя часть каждой ее ветви отведена наружу. Зубная формула такая же, как у опоссумовых: I5/i4, С1/с1, Р3/р3, М4/m4 = 50. При этом верхние и нижние резцы относительно шире и имеют лопаточковидную форму. I1 и i1 отделены друг от друга небольшими диастемами. Нижние резцы расположены на крае челюсти менее скученно, чем у опоссумовых. Клыки небольшие: верхний не доходит до линии, проведенной от кончика Р3 до кончика I1, а нижний немного превосходит по высоте p3. P1 и P2 почти равны по размерам и меньше P3; нижние предкоренные постепенно увеличиваются от p1 до p3. Верхние коренные расположены дугообразно и постепенно сужаются в передне-заднем направлении от M1 до M3. Протокон сравнительно больше, чем у опоссуммовых. Метакон лишь немного больше, чем паракон (за исключением M4). Оба бугорка сдвинуты по направлению к внешнему краю коронки. Отчетливое иетастилярное расширение отсутствует (лишь незначительно развито на M3). Стилярная полка значительно уменьшена и почти одинакова по ширине; стилярные бугорки очень малы. M4 значительно редуцирован (по существу, находится в зачаточном состоянии); его коронка аналогична таковой M1-3, за исключением того, что метакон расположен более медиально. Нижние коренные несколько уменьшаются от m1 до m3. На m1-3 талонид с бассейном и развит лучше тригонида. Тригонид относительно ниже, чем у опоссумовых. 3 тригонидных бугорка крупные, хотя параконид всегда немного меньше, чем метаконид. m4 значительно уменьшен (еще больше, чем у ценолестовых (Caenolestidae)), редукции в наибольшей мере подвергся талонид — узкий и одновершинный. Молочный зубной набор отличается от постоянного наличием дополнительного нижнего резца. По сравнению с другими сумчатыми, для зверков характерно большое число зубных аномалий. Они присутствуют у 8,8 % исследованных образцов и обычно представляют собой лишние зубы и слияния зубов. Щечной аппарат и слизистый язык в совокупности представляют собой эффективный механизм для ловли мелких беспозвоночных. Кишечник сравнительно короткий, что свидетельствует о преобладании в рационе животной составляющей.
В кариотипе 14 хромосом; половые хромосомы — XY/XX.
|
Поиск |