Тапиноцефаловые — Tapinocephalidae
Класс †Синапсиды (Synapsida)
Подкласс †Терапсиды (Therapsida)
Надотряд †Динообразные (Dinomorpha)
Отряд †Диноцефалы (Dinocephalia)
Подотряд †Тапиноцефалиды (Tapinocephalida)
Надсемейство †Тапиноцефалоиды (Tapinocephaloidea)
Семейство †Тапиноцефаловые (Tapinocephalidae)
Тапиноцефаловые (Tapinocephalidae Owen, 1876) — семейство надсемейства тапиноцефалоидов (Tapinocephaloidea Owen, 1876), происходящее из средней перми Африки. Установлены 3 подсемейства с примерно 7-14 родами. Истинное количество родов неизвестно, так как многие из них оказались синонимами, поскольку описывались на основании остатков разнополых и разновозрастных особей одного и того же вида.
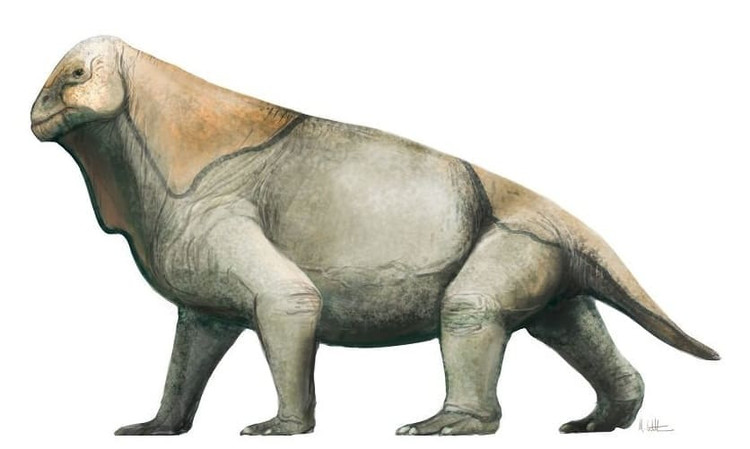

Реконструкция внешнего облика мосхопса (Moschops capensis). Автор — М. Уиттон.
Весьма однородная группа специализированных растительноядных диноцефалов.
Размеры средние или крупные: длина черепа составляет 35-80 см, общая длина варьирует в пределах 1,8-4 м, вес оценивается в 0,3-2 т.
Череп небольшой относительно тела, но очень массивный. Морда удлиненная и низкая, либо умеренно укороченная, высокая и широкая. Дорсальные отростки предчелюстных костей длинные и иногда почти достигают лобных. Ноздри маленькие, низко опущенные. Глазницы небольшие, в большинстве случаев расположенные посредине черепа. Лобные кости часто исключены из ограничения глазниц. Заглазничная дуга массивная, обычно широкая и утолщенная. Чешуйчатая кость достигает заглазничной. Квадратно-скуловая кость часто уменьшена. В результате мощного развития окружающих костей височное окно заметно редуцировано, иногда щелевидное, резко укороченное в передне-заднем направлении и лишенное антеромедиальных вырезок в теменной области, открытое сверху. Книзу чешуйчатая кость очень высокая и оттесняет скуловую кость от края височного окна. Теменная кость обычно принимает участие в образовании его верхнего края. Мозговой отдел короткий, высокий и широкий. Пинеальное отверстие значительно смещено вперед от края затылка.

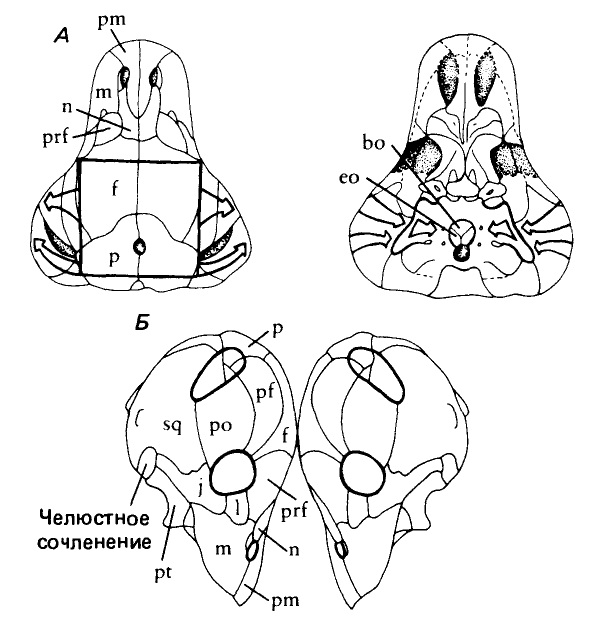
Черепа тапиноцефала (Tapinocephalus atherstonei) (слева) и струтиоцефала (Struthiocephalus whaitsii) (справа).
Надглазничная и лобно-теменная области почти всегда несут сильные пахиостозные разрастания, так что толщина черепной крыши иногда достигает 30 см. Внутричерепные структуры также очень прочные. Высказано предположение, что черепные утолщения использовались для внутривидового соперничества — "бодания", предполагаемого также для пахицефалозаврид и наблюдаемого ныне у козлов и баранов. Противники этой гипотезы указывают, что никаких следов ороговения кожи на лбу и темени не обнаружено. С другой стороны, составляющие их кости обильно пронизаны каналами кровеносных сосудов, вполне могли быть покрыты относительно мягкой кожей и служить для отвода лишнего тепла.
Как правило, затылок очень широкий. Он слабо вогнут и сильно наклонен верхней частью назад. Дистальные концы околозатылочных отростков заднеушной кости отогнуты вниз. Затылочный мыщелок отогнут вентрально или постеро-вентрально. Передняя часть черепа в естественном положении была в той или иной степени опущена вниз. Благодаря особенностям сочленения черепа с позвоночником голова обладала специфической возможностью нагибаться.

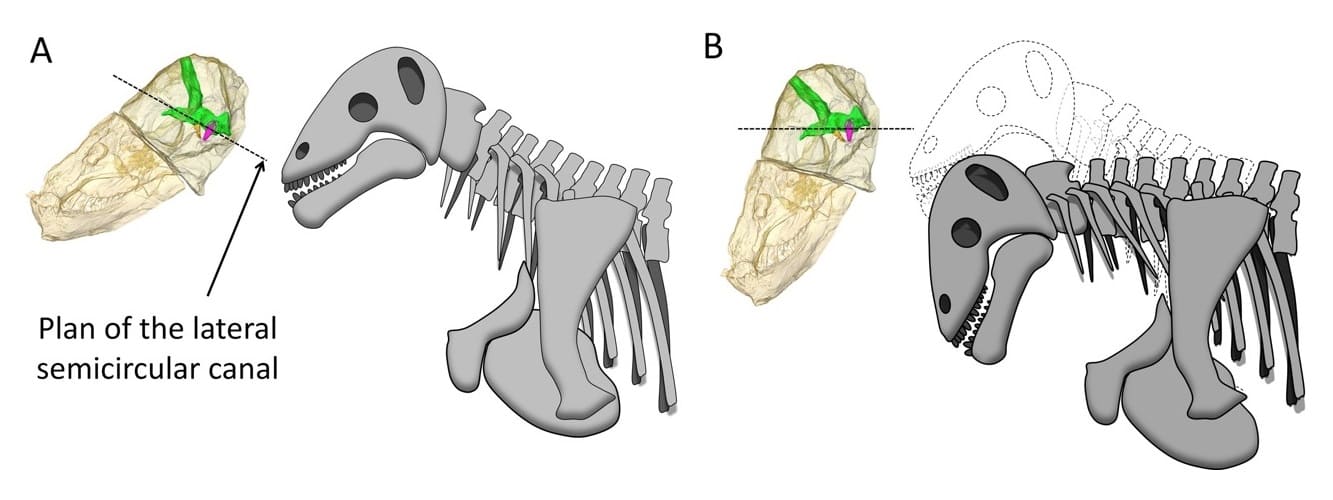
Череп с нижней челюстью мосхопса (Moschops capensis), его компьютерные томограммы и схемы возможного положения головы — традиционно реконструируемая (А) и реконструированная исходя из положения внутреннего уха (В). Зеленым цветом на томограммах выделена полость головного мозга, фиолетовым — внутреннее ухо, служившее органом слуха и равновесия. Судя по ориентации последнего, животное держало голову мордой вниз.
Зубной край предчелюстных костей слабо скошен вверх. Крыловидные кости почти всегда редуцированы и лишь немного заходят за уровень поперечных флангов. Последние сравнительно небольшие и низкие. Крыловидные кости соединяются по срединной линии непосредственно спереди базисфеноида, но впереди их контакта на уровне поперечных флангов птеригоидов имеется хорошо развитая межптеригоидная щель. Базиптеригоидные отростки укорочены. Небные зубы сильно редуцированы или полностью отсутствуют. В предушной области в стенке мозговой коробки иногда хорошо окостеневает латеросфеноид. Сфенэтмоид разрастается назад и может соединяться с переднеушной костью. Слуховая кость обычно прободена отверстием. Овальное окно слуховой капсулы смещено вентрально. Челюстное сочленение, как и у Titanosuchidae, смещено далеко вперед и вниз от затылочного. Нижняя челюсть высокая, массивная, реже тонкая, сравнительно слабая, обычно с ровным альвеолярным краем.
Укороченному и высокому черепу соответствовала сильно развитая и продвинутая в заглазничную область мускулатура, что существенно усиливало давление на зубы передней части черепа.
А — прорисовки реконструированного черепа мосхопса (Moschops capensis). Квадратом выделена основная область утолщения костей крыши черепа, стрелками показано предполагаемое распределение сил, возникающих при бодании головой. Б — ориентация черепов при гипотетическом столкновении лбами.
Зубы относительно небольшие, гомодонтные. В предчелюстной кости имеется 3-5 зубов, в верхнечелюстной — 8-17, в нижней челюсти — 14-19. Особо примечательны передние — долотообразные резцы с хорошо развитыми пяточными выступами и гребнями на лингвальных поверхностях. Как правило, они гораздо крупнее остальных, в связи с чем передние концы предчелюстных и зубных костей очень массивны. Сильно развитые площадки стирания, в виде выпуклости на верхних и соответственного вдавления на нижних резцах, могут свидетельствовать о примитивном передавливании пищи во рту, без перетирания (необходимо отметить, что у большинства растительноядных позвоночных для измельчения служат не передние, а задние, заклыковые зубы). Клыки не дифференцированы, по форме и функционально полностью уподоблены резцам. Увеличенные в числе заклыковые зубы мелкие, колышкообразные, также с пятками и внутренними гребнями, но с меньше выраженными поверхностями стирания; вероятно, они служили в первую очередь для удержания пищевых масс.
Такая зубная система, с сильно удлиненными резцами, была предназначена для захватывания не жесткого, но достаточно плотного растительного корма. Отсутствие структур вторичного неба говорит об очень кратковременном пребывании пищевых масс в ротовой полости — по принципу "схватил и проглотил". Как и у других растительноядных, значительные размеры тела способствовали увеличению длины пищеварительного тракта.
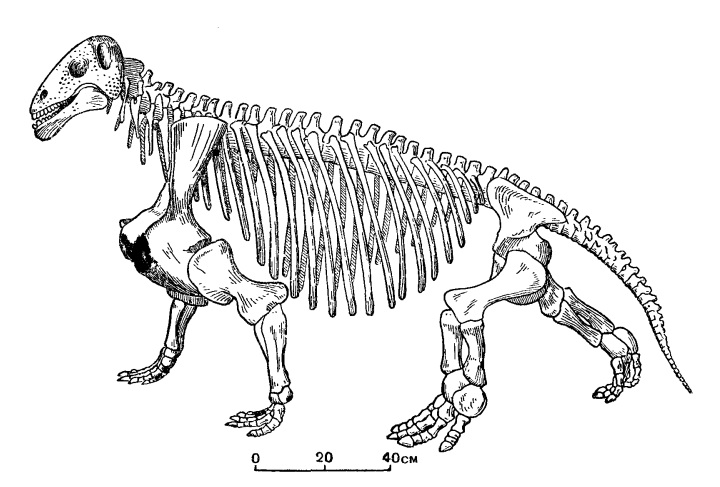
Прорисовка реконструированного скелета мосхопса (Moschops capensis).
 Следовая дорожка тапиноцефалида из средней перми Гансфонтейна в Западной Капской провинции, ЮАР.
Следовая дорожка тапиноцефалида из средней перми Гансфонтейна в Западной Капской провинции, ЮАР.
Шкура тапиноцефалид, вероятно, была мягкой и лишенной чешуй, с многочисленными железами. На лицевой части черепа у некоторых видов обнаружены желобчатые углубления, возможно, вмещавшие кожные железы. Очевидно, для этих животных была свойственна инерционная гомойотермность, их объемистое туловище и короткий хвост как нельзя лучше подходят для сохранения вырабатываемого организмом тепла.

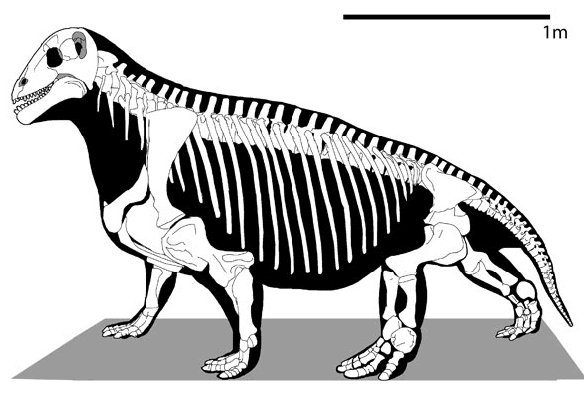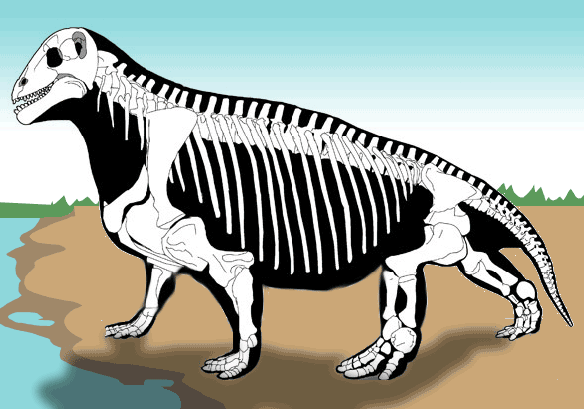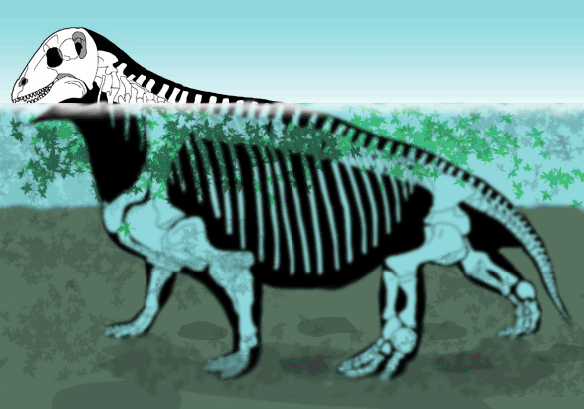

Посткраниальный скелет тяжелый, с относительно хорошо окостеневшими эпифизарными частями. Некрупная голова высоко приподнята на толстой шее. Туловище тяжелое и округлое, приподнятое в своей передней части, так что спина своеобразно поката назад. Плечевой пояс умеренно массивный. Лопатка невысокая, со слабо вогнутым задним краем. Передний отдел межключицы расширен незначительно. Прокоракоид обычно не участвует в ограничении гленоидной впадины. Сохраняется клейтрум. В ацетабулярной впадине и симфизе таза окостенение обычно слабое. Верхняя пластина подвздошной кости невысокая, с удлиненными передним и задним крыльями. Конечности чрезвычайно мощные, передние несколько длиннее и массивнее задних. Передняя их пара сохраняет "растопыренное" положение, но задняя ориентирована почти вертикально. Плечевая кость без эктепикондилярного отверстия. Ступни короткие и широкие. Фаланговая формула маммального типа: 2 - 3 - 3 - 3 - 3. Концевые фаланги копытцеобразно расширены. Хвост короткий.

Шкура тапиноцефалид, вероятно, была мягкой и лишенной чешуй, с многочисленными железами. На лицевой части черепа у некоторых видов обнаружены желобчатые углубления, возможно, вмещавшие кожные железы. Очевидно, для этих животных была свойственна инерционная гомойотермность, их объемистое туловище и короткий хвост как нельзя лучше подходят для сохранения вырабатываемого организмом тепла.
Больше всего тапиноцефалид обнаружено в Южной Африке. Все они характеризуют различные горизонты зоны Tapinocephalus (кептенский ярус, конец средней перми) формации Карру и вне ее не встречаются. Кроме того, имеются находки представителей группы из близких по возрасту слоев формации Рухуху в Танзании.
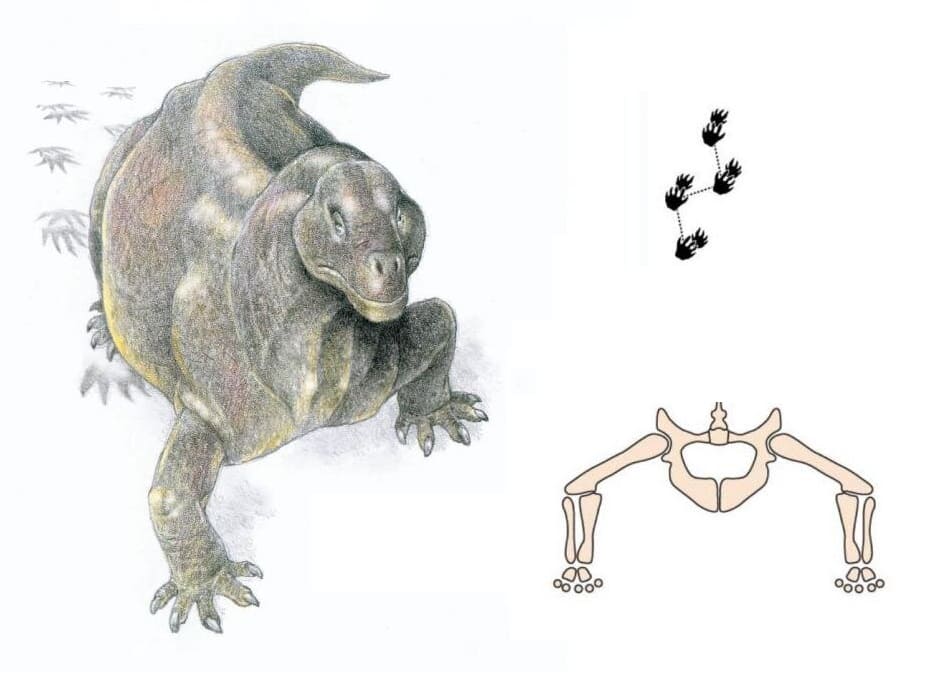
Тапиноцефалид, оставляющий следовые отпечатки, их прорисовка и схема постановки передних конечностей.
Тапиноцефалиды образуют широкий спектр громоздких, бегемотоподобных форм — по-видимому, прибрежных и факультативно-водных. Своими округлыми очертаниями они не похожи ни на одно из современных животных. Наклонная постановка высоко расположенной головы и ее способность нагибаться скорее всего связана с питанием. Видимо, речь идет о поедании каких-то растительных кормов, находившихся ниже уровня головы и не требующих срезания или срывания, поскольку зубная система для этого не приспособлена. Основу рациона, например, могли составлять мертвые стволы каламитов, накапливавшиеся в мелких заводях и лагунах, а также на их берегах. Также существует гипотеза об использовании в пищу плотных, сбитых волнами, полуразложившихся, подвергнутых ферментации и прорастанию микромицетами отмерших масс растений (видимо, в основном представленных членистостебельными) в мелких, солоноватоводных и хорошо прогреваемых прибрежных частях водоемов. Такие маты, состоящие из стволов членистостебельных, известны для соответствующих отложений в ископаемом состоянии.
Схемы, демонстрирующие возможность питания мосхопса (Moschops capensis) взвешенной в воде растительностью.
Судя по всему, тапиноцефалиды представляют собой особое направление эволюционного развития диноцефалов, от относительно примитивных Riebeeckosaurinae (височная область черепа относительно удлинена), через Tapinocephalinae (череп относительно высокий и укороченный) к Struthiocephalinae (наиболее сильно выражена клинориния и удлинена опущенная предглазничная часть).
Несмотря на эволюционную успешность и относительное разнообразие, тапиноцефалиды просуществовали довольно непродолжительное время. Причины их исчезновения до конца не выяснены. Представляется вероятным, что к этому привел не какой-либо один фактор, а неблагоприятное стечение нескольких обстоятельств, например, климатические пертурбации и изменения растительности.

Соперничающие тапиноцефалиды. Автор — Д'Оливейра.
Семейство имеет следующий состав:
подсемейство †Риебеекозаврины (Riebeeckosaurinae Boonstra, 1952);
род †Риебеекозавры (Riebeeckosaurus Boonstra, 1952 );
подсемейство †Тапиноцефалины (Tapinocephalinae Owen, 1876);
триба †Мосхопини (Moschopini);
род †Криоцефалозавры (Criocephalosaurus Kammerer et Sidor, 2002);
род †Мосхопсы (Moschops Broom, 1911);
? род †Авенантии (Avenantia Boonstra, 1952);
? род †Дельфиногнаты (Delphinognathus Seeley, 1928);
род †Мосхопсы (Moschops Broom, 1911);
триба †Тапиноцефалини (Tapinocephalini);
род †Тапиноцефалы (Tapinocephalus Owen, 1876);
? род †Фокозавры (Phocosaurus Seeley, 1888);
? род †Мормозавры (Mormosaurus Watson, 1914);
род †Кератоцефалы (Keratocephalus von Huene, 1931);
подсемейство †Струтиоцефалины (Struthiocephalinae Boonstra, 1969);
род †Струтиоцефалы (Struthiocephalus Haughton, 1915);
? род †Струтиоцефалоиды (Struthiocephaloides Boonstra, 1952);
? род †Струтионопсы (Struthionops Boonstra, 1952);
? род †Тавроцефалы (Taurocephalus Broom, 1928).
Ссылки
Тапиноцефалы (Википедия) перейти
Tapinocephalidae (Paleobiology Database) перейти
Литература
Богданов Д. В. Звероящеры и другие пермские монстры. — М.: Фитон XXI, 2018
Ивахненко М. Ф. Тетраподы Восточно-Европейского плаката — позднепалеозойского территориально-природного комплекса. — Пермь, 2001
Ивахненко М. Ф., Голубев В. К., Губин Ю. М., Каландадзе Н. Н., Новиков И. В., Сенников А. Г., Иванов А. О., Черепанов Г. О. Ископаемые высшие позвоночные. — СПб.: Издательство Санкт-Петербургского университета, 2001
Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Ч. 1 / Отв. ред. Ивахненко М. Ф. и Курочкин Е. Н. — М.: ГЕОС, 2008
Кэрролл Р. Палеонтология и эволюция позвоночных. В 3 т. Т. 2 — М.: Мир, 1993
Основы палеонтологии. В 15 т. Т. 12: Земноводные, пресмыкающиеся, птицы / Гл. ред. Орлов Ю. А. — М.: Наука, 1964
Ромер А. Ш. Палеонтология позвоночных. — М. — Л.: Государственное научно-техническое издательство нефтяной и горно-топливной литературы, 1939
Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М.: ГЕОС, 2009
Чудинов П. К. Ранние терапсиды. — М.: Наука, 1983